Article
한국재래닭에 있어 조우성과 만우성 깃털의 발생 양상 및 형태적 특성 고찰
방민희1, 조은정2, 조창연3, 손시환4,†
Study on the Characteristics of Feather Developing Pattern and Morphology in Early- and Late-Feathering Korean Native Chickens
Min Hee Bang1, Eun Jung Cho2, Chang Yeon Cho3, Sea Hwan Sohn4,†
Author Information & Copyright ▼
1Student, Department of Animal Science and Biotechnology, Gyeongnam National University of Science and Technology,
2inju 52725, Republic of Korea
3Researcher, Department of Animal Science and Biotechnology, Gyeongnam National University of Science and Technology,
4inju 52725, Republic of Korea
5Researcher, Animal Genetic Resources Research Center, National Institute of Animal Science, RDA, Namwon 55717, Republic of Korea
6Professor, Department of Animal Science and Biotechnology, Gyeongnam National University of Science and Technology,
7inju 52725, Republic of Korea
ⓒ Copyright 2018, Korean Society of Poultry Science. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Received: Jun 14, 2018 ; Revised: Aug 05, 2018 ; Accepted: Aug 09, 2018
Published Online: Sep 30, 2018
적요
닭의 깃털은 날개깃과 꼬리깃의 발달과 형태에 따라 조우성과 만우성으로 분류할 수 있다. 현재, 병아리 성 감별법 중 조우성과 만우성을 이용하여 부화 시 깃털 발생양상의 차이에 따른 깃털감별법이 산업적으로 가장 널리 이용되고 있다. 본 연구는 한국재래닭에 있어 자가성감별 계통 조성을 위하여 조우성 병아리와 만우성 병아리의 분류 방법을 제시하고자 한 것으로 한국재래닭 856수를 대상으로 발생 시부터 55일령까지 병아리의 날개깃과 꼬리깃의 발달 양상과 형태를 분석하였다. 또한 K-특이 유전자 프라이머를 이용한 PCR로서 조우성과 만우성 닭의 표현형과 유전자형 간의 일치도를 확인하였다. 분석 결과, 조우성 병아리는 긴 주익우와 부익우를 가지며, 주익우와 부익우 간 확실한 길이의 차이를 보였다. 반면, 만우성 병아리는 조우성 병아리보다 짧은 주익우와 부익우를 보였으며, 주익우와 부익우 간 길이의 차이는 거의 없는 것으로 나타났다. 만우성 병아리는 날개깃 모양에 따라 LF-Less, LF-Scant, LF-Equal 및 LF-Reverse와 같은 4가지 유형으로 분류할 수 있었다. 조우성 병아리의 주익우는 만우성 병아리에 비해 15일령까지 1.5배 정도 더 길다가 50일령에 거의 비슷하게 나타났다. 꼬리깃의 경우, 조우성 병아리는 5일령 때 명확하게 보이지만, 동일시기에 만우성 병아리는 짧고 불명확하였다. 발생 시 주익우 길이 9 mm를 기준하여 조우성과 만우성 개체로 구분하였을 때 조우성은 96.2%, 만우성은 85.4%의 분류의 정확도를 나타내었다. 결론적으로 한국재래닭에 있어 조우성 병아리와 만우성 병아리의 깃털 형태 및 발달 양상의 차이가 뚜렷하기 때문에 발생 시점에서 조우성과 만우성을 쉽게 구분할 수 있을 것으로 판단된다.
ABSTRACT
Chicken feathers could be classified into early-feathering (EF) and late-feathering (LF) depending on the development and patterns of the wing and tail feathers. Currently, feather-sexing is a widely used chick sexing method in the industry. This study was carried out to suggest the method of classifying of EF and LF chicks to establish auto-sexing Korean native chicken (KNC) strains. The development and morphology of wing feathers and tail feathers in 856 KNCs from hatching to 55-days old were analyzed to classify EF and LF chicks. We also performed PCR analysis using K-specific gene primers to confirm the agreement between the phenotypes and genotypes of EF and LF chickens. In the results, the EF chicks had long primaries and coverts, and there was a significant difference in length between primaries and coverts. The LF chicks had shorter primaries and coverts than the EF chicks, and showed little difference in the length between primaries and coverts. LF chicks could be classified into four groups: LF-Less, LF-Scant, LF-Equal and LF-Reverse according to their wing feather patterns. EF chicks had 1.5 times longer primaries than LF chicks until they were 15-days old, but the lengths were almost the same at 50-days old. The tail feathers of the EF chicks were apparent at 5-days old, but those of the LF chicks were short and indefinite at that time. When EF and LF chicks were classified by the length of primaries being more or less than 9 mm, the classification accuracies for EF and LF chicks were 96.2% and 85.4%, respectively, compared to the PCR results. In conclusion, juvenile EF and LF KNC showed distinct differences in feather development and morphology, and could be easily distinguished at one day-old.
Keywords: 한국재래닭; 조우성 병아리; 만우성 병아리; 깃털분류; 깃털 성 감별
Keywords: Korean native chicken; early-feathering chick; late-feathering chick; feather classification; feather-sexing
서 론
발생 직후 병아리의 암수 감별은 산업적으로 매우 중요하다. 그러나 병아리의 해부 생리적 특성상 발생 직후 외관상으로 암수의 구분은 거의 불가능하다. 따라서 발생 병아리의 성의 감별을 위하여 다양한 방법들이 개발되어 소개되고 있는데, 대표적으로 항문돌기감별법(Masui et al., 1925; Martin, 1934), 기계감별법 및 반성유전을 이용한 깃털 성 감별법 등이 있다(Mueller and Moultrie, 1952; Plumart and Mueller, 1954; Siegel et al., 1957; Somes, 1969; Warren, 1976; McGibbon, 1977; SAFRS, 2011). 최근에 분자생물학적 기법을 이용한 병아리의 성 식별이나(Saitoh and Mizuno, 1992; Klein and Ellendorff, 2000; Sohn et al., 2012), 부화 전 수정란 상태에서 호르몬을 이용한 성의 감별(Weissmann et al., 2013) 및 근적외선 형광 라만 분광법(Fluorescence and Raman spectroscopy)을 이용하여 초기 배자 혈액으로부터 성 감별(Galli et al., 2018) 등 다양한 첨단 기법들이 소개되고 있으나, 이러한 방법들이 아직 산업적으로 적용되기에는 많은 어려움이 있다.
이러한 병아리 성 감별법 중 반성유전을 이용한 성 감별이 상업적으로 실용화되고 있는데, 특히 닭의 깃털 발생속도와 연관된 깃털 조만성을 이용한 감별법이 가장 널리 이용되고 있다. 이는 닭의 깃털 발생속도에 관여하는 만우성 특이 유전자인 K가 Avian ev-21 gene 삽입부위 또는 프로락틴 리셉터 유전자인 PRLR과 정자편모단백질 유전자인 SPEC2와 연관하여 Z 염색체에 위치함으로 조우성과 만우성의 반성유전현상을 이용하여 발생 병아리의 성을 식별할 수 있는 원리이다(Humphries et al., 1984; Bacon et al., 1988; Lakshmanan et al., 1992; Iraqi et al., 1995; Bitgood, 1999; Elferink et al., 2008; Zhao et al., 2016), 즉, 종계의 모계를 만우성(ZKW), 부계를 조우성(ZkZk)으로 하였을 때 생산되는 암평아리는 조우성(ZkW)이 되고, 수평아리는 만우성(ZKZk)이 됨으로 깃털 형태로서 성을 판별할 수 있는 방법이다. 이러한 깃털 성 감별법은 다른 성 감별법에 비해 여러 가지 장점이 있는데, 특별한 기술 없이 병아리의 깃털 형태로서 쉽고 빠르게 감별할 수 있으며, 감별에 따른 소요 비용이 발생하지 않기 때문이다. 그러나 깃털의 조만성을 이용하여 생산되는 자손의 암수 감별이 가능하기 위해서는 반드시 부계는 조우성, 모계는 만우성으로 계통 조성이 이루어져야 하고, 이들 종계들의 생산능력도 검증되어야 한다(Saeki and Katsuragi, 1961; Goodman and Muir, 1965; Lowe and Garwood, 1981; Harris et al., 1984; Fotsa et al., 2001; Khosravinia, 2009; Sohn et al., 2013). 현재 대부분의 다국적 육종회사에서는 조우성과 만우성 형질을 도입한 자가성감별 계통을 조성하고, 병아리의 날개 깃을 이용한 성 감별을 실용화하고 있지만, 품종 및 계통에 따라 조우성과 만우성 개체들의 깃털 형태가 다양하여 이를 명확히 구분하는데 다소의 어려움이 있다(Siegel et al., 1957; SAFRS, 2011; Sohn et al., 2012). 조우성 깃털과 만우성 깃털의 구분에 있어서 Siegel et al.(1957)은 Rhode Island에서 발생 때 날개 깃 길이와 12일령의 꼬리 깃 발달양상, 3주령 및 5주령에 병아리의 등 깃털 분포 양상을 비교 분석하여 조우성과 만우성 깃털의 발생 형태의 차이가 있음을 확인하였으나, 조우성과 만우성 개체를 구분할 수 있는 명확한 기준을 제시하지는 못 하였다. 한편, Aviagen과 같은 다국적 종계 회사에서 실용 브로일러의 암수감별을 위한 조만우성의 날개 깃 식별 기준은 조우성은 부익우가 주익우에 비해 1/2∼3/4의 길이로 발생하며, 만우성은 주익우와 부익우의 길이가 동일하거나, 주익우보다 부익우의 길이가 길게 발달한다고 제시하였다(Aviagen Brand, 2018). Sohn et al.(2012)도 조우성의 날개 깃은 주익우와 부익우의 길이 차이가 뚜렷하며, 만우성은 주익우와 부익우의 길이 차이가 나지 않는다고 보고하였다. 그러나 실제 현장에서 날개 깃의 양상은 매우 다양하게 나타나, 기존의 기준으로서 조우성과 만우성 개체를 구분하는데 많은 어려움이 있다. 따라서 조우성과 만우성 개체의 구분을 위해서는 보다 구체적이고 명확한 깃털의 형태적 식별 기준이 필요하며, 특히 국내 토종닭의 깃털 자가성감별 계통 조성을 위해서는 한국재래닭에 대한 조우성과 만우성 개체들의 깃털 발육 양상에 대한 구명이 필요하다. 그러므로 본 연구는 국내 토종닭 병아리의 날개 깃 및 꼬리 깃의 발달 양상과 형태를 분석하여 조우성과 만우성 깃털의 형태적 특징을 비교하고, 깃털 발생 형태에 따른 조우성과 만우성 개체의 식별 기준을 제시하고자 하였다.
재료 및 방법
1. 공시동물 및 사양관리
본 시험에 공시된 닭들은 한협㈜이 보유한 토종 종계들을 이용하여 이들 간 교잡으로 생산한 11개 조합의 실용토종닭들로서 총 856수를 대상으로 하였고, 깃털 조만성 분석 결과 이들 중 조우성이 744수이고, 만우성이 112수였다. 시험계는 발생 직후 경남과학기술대학교 종합농장으로 이송하여 12주간 사육하였는데, 모든 병아리들은 강제 환기 및 자동온도조절시스템이 완비된 무창 계사 형태의 육추사에서 3단 3열 배터리형 케이지(74 cm × 60 cm × 35 cm/cage)에서 사육하였고, 1주령 이후 시험 종료 시까지 케이지 당 사육수수는 12∼15수로 하였다. 사료 급이, 점등관리 및 백신 접종 등 일반 사양관리는 본 대학 육계 사육관리 지침에 따라 이루어졌고, 시험에 관련된 닭의 관리 및 취급은 본 대학 동물실험윤리위원회(IACUC 승인번호; 2018-3)의 규정을 준수하여 시행하였다.
2. 깃털 발생 양상 및 형태 분석
병아리의 날개 깃 발생 양상의 확인은 왼쪽 날개깃에서 2번째 주익우(primaries)와 부익우(coverts)의 길이 및 형태를 측정하였다. 조우성과 만우성 개체의 날개 깃 조사는 부화 직후부터 5일령, 7일령, 10일령, 15일령에 관측 측정하였고, 이후 55일령까지는 5일 간격으로 확인하였다. 꼬리 깃은 부화 직후부터 15일령까지 발생 양상을 확인하고, 이의 길이 및 꼬리 깃의 개수를 조사하였다.
3. 조우성과 만우성 개체 식별을 위한 분자생물학적 검정
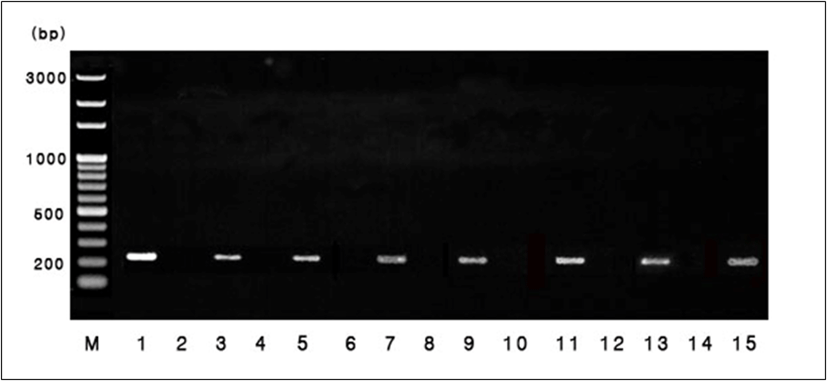
조우성과 만우성 개체 식별을 위한 분자생물학적 검증을 위하여 7주령에 혈액을 채취하고, 이들로부터 genomic DNA를 추출 후 polymerase chain reaction(PCR)을 실시하였다. 조우성과 만우성을 식별하기 위해 사용한 만우성 특이 primer는 ev21 연관 유전자인 JFIL-1의 일부 영역을 이용하여 5’GGGGTCAGCATGTTTAAAGG 3’(forward) 및 5’TTGAGTCCCTAACGATTGCG 3’(reverse)으로 제작하였다(Iraqi and Smith, 1994). 제작된 primer를 이용한 PCR은 TaKaRa TaqTM kit(Takara, Kyoto, Japan)를 사용하였고, PCR 반응물은 10× buffer 2.5 μL, dNTP 2 μL, Taq polymerase 2 μL (0.5 unit/μL), primer 4 μL(5 pmol/μL), 추출한 genomic DNA 3 μL (100 ng/μL) 및 ddH2O 11.5 μL를 넣어 총량이 25 μL가 되도록 혼합하였다. PCR 증폭 반응 조건은 95℃ 5분간 변성시키고, 95℃ 1분간 변성, 56℃ 1분 접합, 72℃ 2분간 신장하는 3단계 과정을 30회 반복 후 72℃에서 15분간 신장시켰다. 증폭된 PCR 산물은 200 ng/μL로 정량하여 전기영동 후 band의 유무에 따라 조우성과 만우성을 확인하였다 (Fig. 1).
Fig. 1.
The results from polymerase chain reaction using the K-specific primer in chickens. M is the 100bp size marker.
Lanes of 1, 3, 5, 7, 9, 11, 13, and 15 are late-feathering chicks. Lanes of 2, 4, 6, 8, 10, 12, and 14 are early-feathering chicks.
Download Original Figure
4. 통계분석
조우성과 만우성 개체의 날개 깃 측정치 평균 간의 차이에 대한 비교 분석은 SAS 통계 패키지(SAS Institute Inc., Cary, NC, USA)를 이용하여 t-test로 평균 간의 유의성을 검정하였다.
결과 및 고찰
1. 발생 직후 병아리 날개깃의 형태 분석
깃털의 형태적 양상으로 구분한 조우성과 만우성 개체의 확인을 위하여 Gallus gallus endogenous virus-21JFIL-1 DNA의 일부영역인 만우성 특이 유전자를 이용하여 PCR 분석을 수행하였다. 분석 결과, Table 1에 제시된 바와 같이 공시된 856수 중 조우성이 746수, 만우성이 110수로 판별되어 깃털의 형태적 특성으로 분류한 개체 수와 단 두 마리의 오차를 보여 99.8%의 일치도를 나타내었다. 따라서 본 연구에서 제시한 깃털 형태에 의한 조우성과 만우성의 판별 기준은 토종닭에서 거의 100% 신뢰할 수 있는 깃털 분류 지표라 사료된다.
Table 1.
The conformity between morphological feathering classification and PCR analysis for the identification of early- and late-feathering chicks
| Feathering types |
Phenotypic analysis |
PCR analysis |
Conformity(%) |
| Early feathering |
744 |
746 |
99.7 |
| Late feathering |
112 |
110 |
98.2 |
| Total |
856 |
856 |
99.8(854/856) |
Download Excel Table
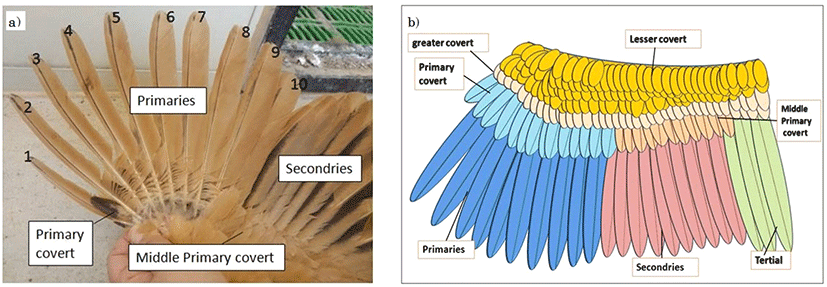
닭의 날개깃은 Fig. 2에 제시된 바와 같이 크게 1차 날개깃(primaries; 주익우)과 날개덮깃(primary covert; 부익우) 및 2차 날개깃(secondaries)과 이의 날개덮깃(middle primary covert) 등으로 이루어져 있으며, 주익우의 개수는 평균 10개 정도였다. 날개깃을 이용한 조우성과 만우성의 분석은 주익우와 부익우를 대상으로 하였는데, 첫 번째 주익우는 깃의 발생이 불완전하거나 손상이 많아 두 번째 주익우와 부익우를 조사 분석하였다. Fig. 3은 한국토종닭 조우성과 만우성 병아리의 발생 직후 날개깃의 형태이다. 조우성의 깃털 형태는 주익우와 부익우 간에 뚜렷한 길이의 차이를 보이고, 부익우는 주익우의 1/2∼1/3 정도의 길이를 나타내었다. 이들의 깃털 형태에 있어 깃가지의 발생이 듬성하고 미숙한 경우 EF-1으로, 깃가지의 발생이 조밀하고 왕성한 경우를 EF-2로 임의 분류하였는데, 두 종류 간 깃가지의 발생 형태만 다를 뿐 주익우와 부익우의 길이나 양상은 동일하였다. 반면, 만우성은 주익우와 부익우 간의 길이와 형태에 따라 임의로 LF-Less, LF-Scant, LF-Equal 및 LF-Reverse의 4가지 유형으로 분류하였다. LF-Less 유형은 주익우와 부익우 간의 길이 차이가 뚜렷하게 나타나지만, 주부익우의 길이가 모두 조우성에 비해 짧다. LF-Scant 유형은 부익우의 길이가 1∼2 mm 정도로 깃의 길이가 매우 짧은 양상이다. LF-Equal 유형은 주익우와 부익우의 길이 차이가 거의 나지 않으며, LF-Reverse 유형은 특이적으로 부익우의 길이가 주익우에 비해 길게 나타나는 형태이다. 본 시험에서 만우성으로 분류된 112개체의 유형별 빈도는 LF-Equal 유형이 52%로 가장 많았으며, LF-Less는 35%, LF-Reverse는 11%로 조사되었고, LF-Scant 유형은 2%로 가장 낮게 나타났다. 조우성 깃털과 만우성 깃털의 유형별 특징에 대하여 Aviagen Brand(2018)는 브로일러의 경우 주익우와 부익우의 길이가 비슷하거나, 부익우가 주익우보다 긴 양상만을 만우성으로 판별하였고, Sohn et al.(2012)은 한국재래닭에서 조우성은 주익우가 부익우에 비해 성장이 빠르고 이들 간 길이의 차이가 뚜렷한 반면, 만우성은 주익우와 부익우의 발생 양상이 비슷하다고 제시하였으나 만우성 깃털의 형태가 다양하여 구분이 모호한 경우가 많다고 하였다. 또한 Somes(1969; 1970)와 McGibbon(1977)도 조만우성의 구분에서 실제 날개깃 형태 및 발달 속도가 매우 다양함을 보고하였다. 따라서 본 연구에서는 Fig. 4와 같이 1일령 병아리의 주익우와 부익우의 길이 및 형태적 양상에 따라 조우성과 만우성 개체들의 유형별 형태를 설정하여 조우성과 만우성의 구분 기준을 제시하였다.
Fig. 2.
Chicken wing feather;
a) Photograph of chicken wing feathers and number of primaries. b) Diagram of chicken wing feather structure based on the left figure.
Download Original Figure
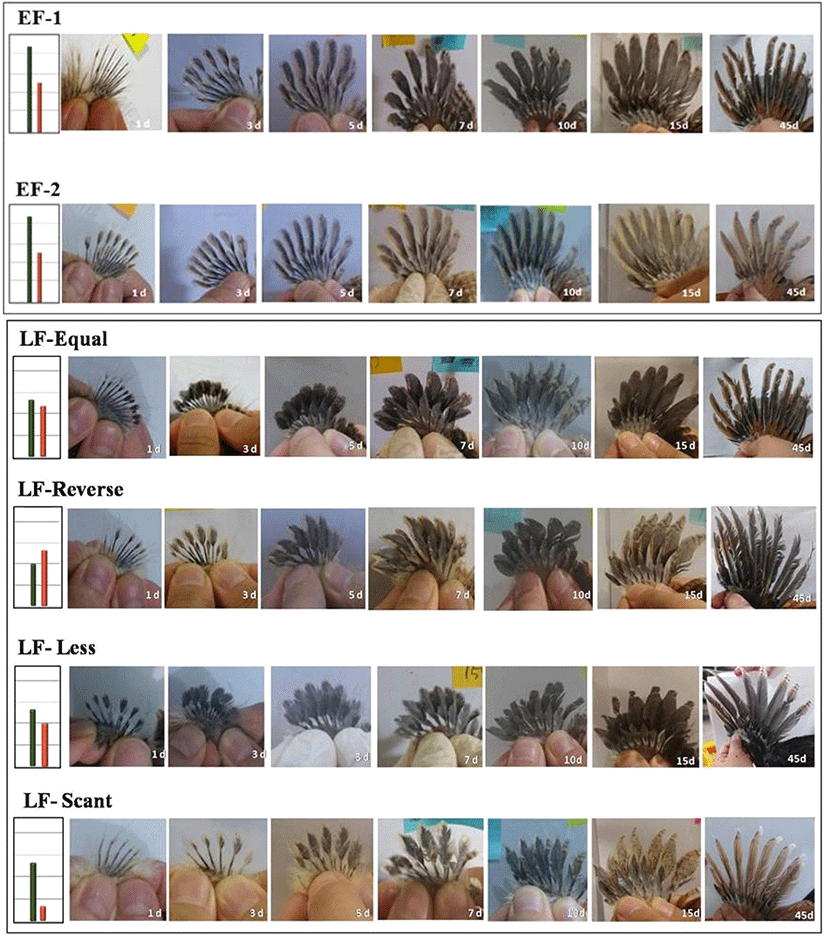
Fig. 3.
The patterns of wing feather in early-feathering and late-feathering day-old chicks.
EF-1 and EF-2 are early-feathering types, and LF-Less, LF-Scant, LF-Equal and LF-Reverse are late-feathering types.
Download Original Figure
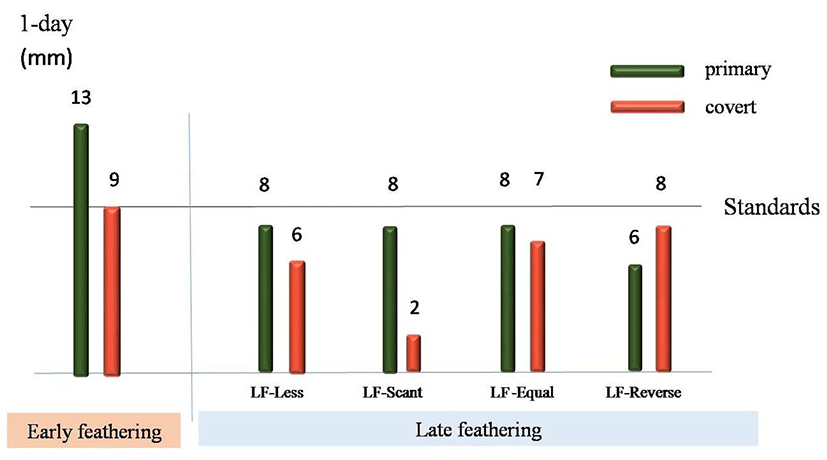
Fig. 4.
The diagram of wing feather types of early-feathering and late-feathering chicks at day-old.
The diagram was based on the length of second primary and it’s covert. LF-Less showed that both of the primaries and coverts were shorter than the early-feathering chick’s. LF-Scant had almost no coverts. LF-Equal had the same length of primaries and coverts. LF-Reverse showed the coverts were longer than the primaries.
Download Original Figure
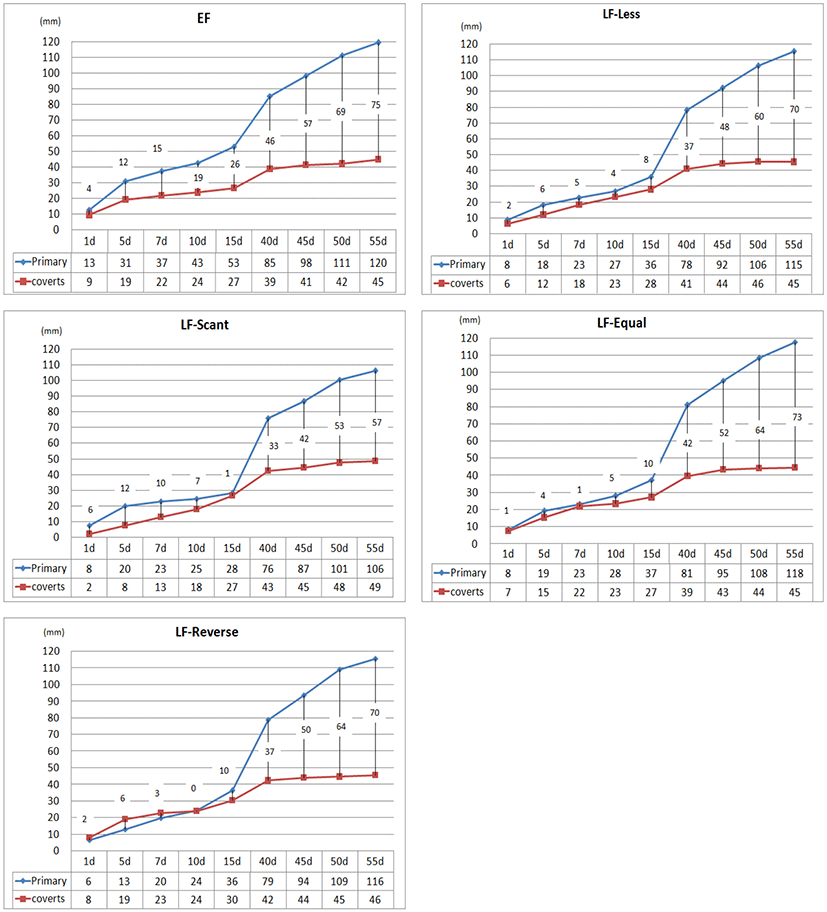
2. 조우성과 만우성 병아리의 날개깃의 발달 양상
조우성과 만우성 병아리의 날개깃의 발달 양상을 조사하기 위하여 부화 직후부터 55일령까지 날개깃의 길이와 형태를 비교 분석하였다. 1일령부터 45일령까지 관찰한 조우성 및 만우성의 날개깃 발달 형태는 Fig. 5와 같고, 일령에 따른 주익우와 부익우 길이의 변화 양상을 Fig. 6에 나타내었다. 본 결과에서 EF-1이나 EF-2의 깃의 발달 양상은 동일하였고, 공히 일령이 증가함에 따라 주익우의 발달이 부익우에 비해 현저히 빠름을 확인하였다. 주익우와 부익우의 길이를 측정한 결과, 1일령의 평균 날개깃의 길이는 주익우가 13 mm, 부익우가 9 mm로 나타났으며, 이 후 이들 간 길이의 차이가 점점 증가하면서 7일령에는 주익우의 길이가 부익우 길이의 1.7배, 15일령에는 2배, 55일령에는 2.6배 정도로 나타났다. 반면, 만우성 개체들의 날개깃의 발달 형태는 유형별로 다소 차이는 있으나, 대부분 15일령까지 주익우와 부익우의 성장 속도가 비슷하며, 이들 간의 길이의 차이는 그리 크지 않은 것 나타났다. LF-Less 유형은 주익우와 부익우의 길이 차이가 나고 성장 양상이 조우성과 비슷하지만, 깃의 길이가 짧고 발달 속도가 전체적으로 늦은 것으로 나타났다. LF-Scant는 발생 때 부익우의 길이가 현저하게 짧은 양상으로 10일령까지 부익우의 발달이 매우 느리고 15일령 이후 주부익우의 성장 양상이 다른 유형과 비슷하게 나타난다. LF-Equal은 발생 때 주익우와 부익우의 길이가 비슷한 형태로 주부익우 간 발달 양상이 7일령까지 거의 동일한 양상이다가 10일령 이후 주익우와 부익우 간 길이의 차이가 나타난다. LF-Reverse는 발생 때 주익우에 비해 부익우의 길이가 긴 양상인데, 7일령까지는 부익우가 주익우 길이보다 길게 나타나고, 10일령 때 거의 동일한 양상을 보이다가 이후 다른 유형과 발달 양상이 동일하였다.
Fig. 5.
The developmental patterns of wing feathers in early-feathering chicks (EF-1, EF-2) and late-feathering chicks (LF-Equal, LF-Reverse, LF-Less, LF-Scant).
Download Original Figure
Fig. 6.
The length of primaries and coverts in early-feathering chicks (EF) and late-feathering chicks (LF-Less, LF-Scant, LF-Equal and LF-Reverse) from one-day-old to 55-day-old.
Download Original Figure
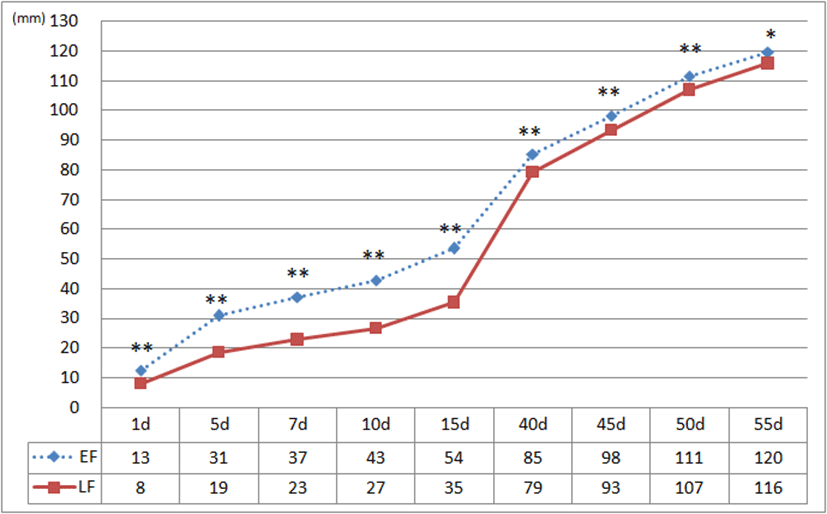
Fig. 7은 조우성과 만우성 병아리의 주익우 길이를 비교 측정한 결과로서 발생 시점부터 55일령까지 모든 일령에서 조우성과 만우성 간에 유의한 차이를 보였다. 어린병아리 때 조우성 개체들은 만우성 개체에 비하여 월등히 긴 주익우를 가지나 15일령 이후부터 이들 간 길이의 차이는 점차적으로 감소하여 40일령쯤에는 깃털의 길이로서 조우성과 만우성을 식별하기는 거의 불가능하였다. 주익우의 길이를 기준으로 조우성이 만우성에 비하여 1일령에는 1.5배, 5일령은 1.7배, 7일령과 10일령은 1.6배, 15일령에서는 1.5배로 나타나고, 15일령 이후부터 이의 길이 차이가 점차적으로 줄어들다가 50일령 쯤에는 거의 동일하였다. 한편, 조우성과 만우성 개체들의 부익우의 길이를 비교 분석한 바, 1일령부터 7일령까지는 조우성이 만우성에 비하여 다소 긴 부익우의 깃 발달을 보였으나, 10일령 이후부터는 이들 간의 차이가 없는 것으로 나타났다. 부익우의 발달 양상은 성장이 진행될수록 조우성과 만우성의 길이가 비슷하였는데, 발생 후 5일령 정도까지만 이들의 길이 차이를 인지할 수 있는 것으로 보여 단순 부익우의 길이만으로 조우성과 만우성 개체의 식별은 어려울 것으로 판단된다.
Fig. 7.
Comparison of primary length between early-feathering and late-feathering chicks from one-day-old to 55-day-old.
* P<0.05, ** P<0.01.
Download Original Figure
이상 국내 토종닭의 병아리를 이용하여 조우성과 만우성의 유형별 양상을 살펴본 바 조우성과 만우성의 날개깃의 발생 양상 및 형태적 차이가 명확히 나타났다. 국내 토종닭의 만우성 깃털은 4가지 유형으로 분류할 수 있었고, 부화 직후인 1일령에서 날개깃의 형태적 차이가 조우성과 만우성 간 뚜렷이 구별되나, 15일령 이후에는 조우성과 만우성 간 거의 유사한 양상으로 발달됨을 알 수 있었다. 발생 직후 1일령 만우성 개체들의 주익우의 평균 길이는 8 mm 정도로서 조우성과 만우성 구분 기준을 주익우 길이를 9 mm로 하여 9 mm 이상을 조우성, 9 mm 미만을 만우성으로 설정했을 때, 조우성의 식별 정확도는 96.2%, 만우성은 85.4%를 보였다. 조우성에 비하여 만우성의 정확도가 낮은 것은 LF-Equal 유형에서 주익우와 부익우 모두 9 mm 이상인 경우가 다수 발생하였기 때문이다. 따라서 발생 직후 주익우의 길이만으로도 어느 정도 조우성과 만우성의 식별이 가능한 것으로 판단된다.
3. 조우성과 만우성 개체의 꼬리깃 발달 양상
병아리의 꼬리깃의 발달양상을 조사하기 위하여 발생 직후 1일령부터 17일령까지 꼬리깃의 발생 유무, 길이 및 개수를 조사하였다. Fig. 8은 조우성 및 만우성 개체들의 꼬리깃의 발달 양상이다. 조우성의 경우 3일령까지 꼬리깃의 발생이 거의 없다가 5일령에는 분명한 꼬리깃의 발달을 나타내었다. 반면, 만우성의 경우 5일령까지 꼬리깃의 발달이 거의 나타나지 않았으며, 대부분의 개체들에서 17일령까지도 꼬리깃의 발달이 명확하지 않았다. Fig. 9는 조우성과 만우성의 꼬리깃의 발생 양상을 좀 더 자세히 비교하기 위하여 5일령부터 17일령까지 병아리 항문을 중심으로 꼬리깃의 형태를 조사한 것이다. 7일령 때 조우성과 만우성의 꼬리깃의 형태를 비교하였을 때 조우성은 꼬리깃의 깃대가 길고, 깃가지가 매우 발달된 양상이었으나, 만우성 개체는 깃대와 깃가지가 짧아 꼬리깃과 솜깃의 구분조차 어려웠다. 이러한 깃 발달 양상은 일령이 증가할수록 명확한 차이를 보이는데, 17일령까지 만우성은 조우성 개체에 비하여 현저하게 발달 속도가 느린 것을 알 수 있었으며, 28일령 이후 1차 환우가 될 때까지 조우성과 만우성 개체 간 꼬리깃의 발달 양상의 차이를 보였다. 이러한 양상은 Siegel et al.(1957)은 조우성과 만우성의 꼬리깃 발생 형태가 12일령쯤 명확한 차이를 보인다고 하였으나, Sohn et al.(2012)은 5일령이면 충분히 구분 가능한 차이를 나타낸다고 하였다. 본 연구 결과에서도 국내 토종닭의 경우, 꼬리깃의 외형적 양상으로 5일령에 조우성과 만우성의 구분이 가능한 것으로 판단된다. 꼬리깃의 수는 조우성과 만우성 공히 8~12개 정도인 것으로 관찰되었으며, 조우성과 만우성 개체 간 개수의 차이는 없는 것으로 나타났다.
Fig. 8.
The development of tail feather of early-feathering chicks (EF) and late-feathering chicks (LF) from one-day-old to 15-day-old.
Download Original Figure
Fig. 9.
The comparison of developmental pattern of tail feathers between the early-feathering chicks and the late-feathering chicks.
Download Original Figure
사사
본 논문은 농촌진흥청 연구사업(세부과제번호: PJ011639 2018)의 지원에 의해 이루어진 것임.
References
Aviagen Brand 2018 Feather sexing day-old chicks in the hatchery. http://en.aviagen.com/assets/Tech_Center/BB_Resources_Tools/AA_How_Tos/AA-How-to-11-FeatherSexDayOldChicks-EN-17.pdf .

Humphries EH, Danhof ML, Hlozanek I 1984 Characterization of endogenous viral loci in five lines of white Leghorn chickens. Virology 135(1):125-138 .

Khosravinia H 2009 Effect of the slow (K) or rapid (k+) feathering gene on carcass related traits of broiler chickens selected for breast and thighs weight. Genetika 45(1):112- 118 .

Lakshmanan N, Ponce de Leon FA, Smyth JR, Smith EJ 1992 Chromosomal assignment of the evil locus in chickens using fluorescent in situ suppression hybridization (FISH). Page 10 In: 10th European Colloquium on Cytogenetics of Domestic Animals. Bosma AA ed. University Press, Utrecht, The Netherlands .
Lowe PC, Garwood VA 1981 Independent effects of K and k+ alleles and maternal origin on mortality and performance of crossbred chickens. Poult Sci 60(6):1123-1126 .
Martin JH 1934 Review the 'sexing baby chicks by Masui and Hashimoto. Poult Sci 13(3):190 .
Masui K, Hashimoto J, Ono I 1925 The rudimental copulatory organ of the male domestic fowl with reference to the sexual differentiation of chickens. Jpn J Zootech Sci 1(3): 3-15 .
McGibbon WH 1977 A sex-linked mutation affecting rate of feathering in chickens. Poult Sci 56(3):872-875 .
Mueller CD, Moultrie F 1952 Classification of sex-linked early and late feathering in 10-week-old chickens. Poult Sci 31(1):171-172 .
Plumart PE, Mueller CD 1954 Effect of sex-linked early feathering on plumage from 6 to 12 weeks of age. Poult Sci 33(4):715-721 .
Saeki Y, Katsuragi T 1961 Effect of early and late feathering gene on growth on New Hampshire, Leghorns and their crossbreds. Poult Sci 40(6):1612-1616 .
SAFRS 2011 Chicken sexing, primary industries and fisheries. Queensland government, Web http://www.dpi.qld.gov. au/27_2712.htm .
Siegel PB, Mueller CD, Craig JV 1957 Some phenotypic differences among homozygous, heterozygous, and hemizygous late feathering chicks. Poult Sci 36(2):232-239 .
Sohn SH, Cho EJ, Kang BS 2012 Sex identification of newly hatched chicks by fluorescence in situ hybridization using a W-specific DNA probe in feather follicle cells. J Poult Sci 49(4):231-236 .
Sohn SH, Kim NY, Park DB, Song HR, Cho EJ, Choi SB, Heo KN, Choi HC 2013 Influence of early- and latefeathering phenotype on productive performance in the feather-sexing strains of Korean native chicken. Korean J Poult Sci 40(3):263-270 .
Sohn SH, Park DB, Song HR, Cho EJ, Kang BS, Suh OS 2012 Genotype frequencies of the sex-linked feathering and their phenotypes in domestic chicken breeds for the establishment of auto-sexing strains. J Anim Sci Tech 54(4):1-10 .
Somes RG 1970 The influence of the rate of feathering allele Kn on various quantitative traits in chickens. Poult Sci 49(5):1251-1256 .
Warren DC 1976 Feather-sexing chicks. Poult Tribune 82(2):32-34 .
Zhao J, Yao J, Li F, Yang Z, Sun Z, Qu L, Wang K, Su Y, Zhang A, Montgomery SA, Geng T, Cui H 2016 Identification of candidate genes for chicken early- and late-feathering. Poult Sci 95(7):1498-1503 .