
서 론
기후변화는 가축 산업이 직면한 가장 도전적 문제 중의 하나가 되었으며, 그 중에서도 열 스트레스는 가축의 생산성뿐만 아니라, 동물복지에도 지대한 영향을 준다. 지구 온난화는 21세기에 접어들면서 더욱 가속화되고 있다. 우리나라(서울 기준)의 여름 지속일은 1910년대(1911∼1920년) 평균 98일에서 2010년대(2011∼2017년) 117일로 19일이 길어졌고, 최근 30년 기온은 20 세기 초(1912∼1941)보다 1.4°C 상승하였다(NIMS, 2018). 길어진 여름과 높은 기온현상은 가축에게 심각한 스트레스로 작용하고, 열 스트레스는 수 시간 혹은 수일을 지속할 수도 있으며, 경우에 따라 수개월을 지속할 수도 있다. 열 스트레스는 가금의 성장률 향상과 높은 생산 효율 등을 기반으로 하는 육종의 결과, 보다 커진 대사량과 체열 생산, 그리고 저하된 열 저항성 등에 의하여 가금 산업에서 특히 더 중요하게 여겨지고 있다(Settar et al., 1999, Deeb and Cahaner, 2002). 따라서 열 스트레스를 완화시키기 위한 다양한 전략들이 요구되는데, 이를 위해서 가축의 열 스트레스에 대한 정확한 평가와 생물학적 평가지표의 개발은 기후변화에 대한 적응과 열 저항성 가축의 육종, 지속 가능한 축산과 동물복지를 위해서 매우 중요하다. 가축의 스트레스에 대한 반응의 정도를 양적으로 나타낼 수 있는 바이오마커에는 corticosterone과 같은 호르몬(Kang and Kuenzel, 2014), 호중구(heterophil)와 림프구(lymphocyte)의 비율(Scanes, 2016), DNA 손상 정도(Sohn et al., 2012), 열충격단백질(heat shock proteins; HSPs) 등이 있다. 이들 바이오마커 중에 HSP는 세포내 온도계로 여겨지며(Katchinski, 2004), HSP 중에서도 HSP70은 가장 풍부한 단백질이면서 열 충격에 민감하게 반응하며, 가축의 온도 적응에 중요한 조절자로서의 기능을 한다(Dangi et al., 2014). 따라서 HSP70 바이오마커는 가축의 열 스트레스에 대한 체계적 평가방법으로 주목되고 있으며, HSP70의 발현 변화의 이해는 기후변화에 대처하면서 지속가능한 가금 산업을 위해 매우 중요하고 판단된다. 이에 본 논문에서는 닭의 열 스트레스와 관련하여 HSP70을 중심으로 HSP의 작용기전과 역할 및 바이오마커로서의 활용 등에 대한 최근까지의 연구들을 소개하고자 한다.
본 론
열충격 단백질은 원핵생물과 진핵생물 모두에서 많이 발현되는 단백질로서 1962년 초파리에서 처음으로 보고되었다(Ritossa, 1996). Ritossa와 연구자들은 초파리를 2∼3분간 열처리(37°C)한 후 염색체의 특정 부위가 부풀어 오르는(puffing) 패턴과 이곳에서 발현되는 유전자들로부터 70과 25 kDa 단백질의 발현 증가를 관찰하였고, 열 충격 실험에 의해 반복적으로 나타난 이들 단백질을 “heat shock protein(열충격단백질)”으로 명명하였다. 열충격 단백질의 분자량은 10∼170 kDa으로 다양하며, 크기에 따라 세포내에서 분포, 기능, 발현 양상 등에 차이가 있다(Snoeckx et al., 2001). 다양한 크기로 발현되는 열충격단백질의 유전자들은 ‘hsp’ genes으로, 이들 유전자들에 의해 번역된 각각의 단백질은 Hsp로, 그리고 Hsp들의 크기별 group(그룹)으로 표현하는 family(계)는 HSP로 표기하기로 결정하여(Hightower and Hendershot, 1997) 본 원고에서도 이에 따라 기술하였다. 열충격단백질들은 분자량이 각기 다른 단백질 군(group)으로 구성되어 있는 단백질 집합체(super family)이다(Table 1). 대표적 예가 HSP70 family로서 분자량 70, 73, 78 kDa 등이 알려져 있다(Snoeckx et al., 2001). 따라서 이들 단백질을 발현하는 유전자를 일반적으로 hsp70이라 부른다. HSPs는 더위와 같은 높은 온도뿐만 아니라, 미생물 감염, 칼슘증가 매개체, 호르몬, 항생제 및 중금속 등과 같은 일련의 스트레스에 노출되었을 때에도 발현된다(Hassan et al., 2019). 열충격 유전자들은 housekeeping gene의 성질을 갖고 있어 세포의 정상적 활동에도 지속적으로 발현이 되지만, 생리적, 환경적 스트레스가 주어지면 발현이 유도되어 새롭게 합성된 HSP는 세포내 필요로 하는 곳으로 빠르게 이동하여 HSPs로서 기능을 하게 된다. HSP중에서 가장 풍부하게 발현되는 HSP70을 종별로 살펴보면, 닭의 Hsp70의 분자량은 69,913Da이며, 634개의 아미노산으로 구성되어 있다. 닭의 Hsp70은 사람(639aa; 유사도 92.5%)을 포함하여, 소(631aa; 유사도 90.3%), 생쥐(633aa; 유사도 91.3%), 쥐(633aa; 유사도 91.2%), 돼지(643aa; 77.3%) 등과 같이 아미노산 유사성이 매우 높다. 진핵생물에서 hsp70의 유전자 염기서열에 대한 유사성은 60∼78%이다(Kiang and Tsokos, 1998). HSP70은 단량체 단백질로 원핵세포의 세포질에 위치하지만, 진핵세포에서는 세포질, 소포체, 핵, 엽록체, 미토콘드리아 등 다양한 곳에 분포하고 있으며, 세포외 기질(extracellular matrix)에서도 발견되고, 반감기는 사람의 상피세포에서 대략 48시간이다(Kiang and Tsokos, 1998).
| HSP family | Family members | Cellular location | Characteristics/functions | Upon stress | References |
|---|---|---|---|---|---|
| HSP27 | Hsp25 Hsp27 |
Cytoplasm/nucleus | Constitutive expression/forms large aggregates | Phosphorylated upon stress/nuclear translocation | Larsen et al., 1995 |
| HSP60 | Hsp58 Hsp60 |
Endoplasmic reticulum/mitochondria | Constitutive expression/guides mitochondrial protein import in conjunction with Hsp70 | - | Snoeckx et al., 2001 |
| HSP70 | Hsp70 Hsp73 Hsp75 Hsp78 |
Cytoplasm/nucleus/mitochondria/endoplasmic reticulum | Constitutive & inducible expression/protein folding /guides protein synthesis and import for protein degradation Hsp70:highly inducible & normally absent | Only Hsp70 inducible at heat stress/guides protein synthesis and binds proteins upon stress/nuclear translocation | Leung et al., 1990. Snoeckx et al., 2001 |
| HSP90 | Hsp90-α Hsp90-β |
Cytoplasm/nuclear membrane/nucleous | Constitutive expression/binds cytoplasmic hormone receptors | Up-regulated and phosphorylated upon stress/nuclear translocation |
Collier NC and Schlesinger, 1986; Snoeckx et al., 2001 |
| HSP104 | Hsp94 Hsp104 Hsp110 |
Cytoplasm/nucleolus | Constitutive expression/protein refolding | Up-regulated upon stress/nuclear translocation |
Hatayama et al., 1994
Snoeckx et al., 2001 |
단백질의 3차구조의 형성에는 단백질의 접힘(folding)이 가장 중요하다. 리보솜에서 새롭게 합성된 폴리펩타이드(polypeptide)가 정확하게 접히기 위해서는 다른 단백질의 도움을 받아야 한다. 폴리펩타이드의 접힘을 도와주는 단백질들을 분자적 샤페론(molecular chaperons)이라 한다. 진핵세포의 경우, 샤페론의 종류와 수가 원핵세포보다 많으며, HSP70은 모든 진핵 및 원핵세포에서 공통적으로 작용한다. HSPs의 종류로는 HSP27, HSP60, HSP70, HSP90, HSP104등이 대표적이다(Table 1). 진핵세포의 조면소포체에서 폴리펩타이드의 접힘에는 HSP70이 주요 샤페론이며, 스테로이드계 호르몬 수용체와 세포신호에 관여하는 단백질들은 HSP90의 추가적 도움을 받는다. HSP단백질은 C말단 도메인을 통해 주변의 변성(denatured), 미접힘(misfolded) 혹은 손상(damaged)된 단백질의 소수성 서열에 결합하여 3차원 구조를 형성(회복)하도록 도와주는 샤페론의 기능을 갖고 있다(Mayer and Bukau, 2005) (Fig. 1). 샤페론의 대표적인 HSP70은 단백질의 정상적 접힘을 통하여 합성된 단백질의 자연적 상태로 유지, 단백질간 응집(일반적으로 독성단백질 복합체 형성) 억제, 응집된 단백질의 재접힘과 용해(해리)성을 높여 세포내 단백질들의 항상성(homeostasis)을 복구하는데 기여한다(Mayer and Bukau, 2005). 이에 더하여 HSP70은 회복되지 못한 문제성 단백질과 기능을 상실한 단백질은 분해가 일어날 수 있도록 돕는 역할도 한다(Yokota and Fujii, 2010). HSP70은 세포내에서 아직 접힙을 하지 않은 단백질들을 안정화시키고, 세포내 단백질의 운송을 촉진한다(Borges and Ramos, 2005). HSPs은 자극에 의해 발현이 유도(유도 단백질: inducible HSPs=iHSPs)되거나 혹은 지속적 발현(constitutively expressed HSPs=cHSPs)을 하는 두 가지 형태가 있으며, 세포질에서 샤페론의 활성이 증가하면서 열 저항성에 도움을 준다(Lindquist and Craig, 1998). 유도형 HSPs는 스트레스에 의해 발현되고, 단백질의 변성, 부적절한 단백질의 축적 등을 방지함으로써 세포의 생존을 강화시켜 준다. HSP에 의한 세포보호 기능은 소장, 신장, 배아 등을 포함한 동물의 여러 장기에서 관찰된다(Baht et al., 2016). 이상의 내용을 종합해 보면 HSP70은 분자적 샤페론으로서 새롭게 합성된 폴리펩타이드의 접힘을 유도하고, 정상적 구조를 획득한 단백질은 세포내 소기관 및 세포밖으로 수송하는 등 단백질의 품질관리에서부터 문제성 단백질을 분해하는 기능에 이르기까지 세포의 정상적 기능과 생존에 필수적인 단백질이라 할 수 있다.
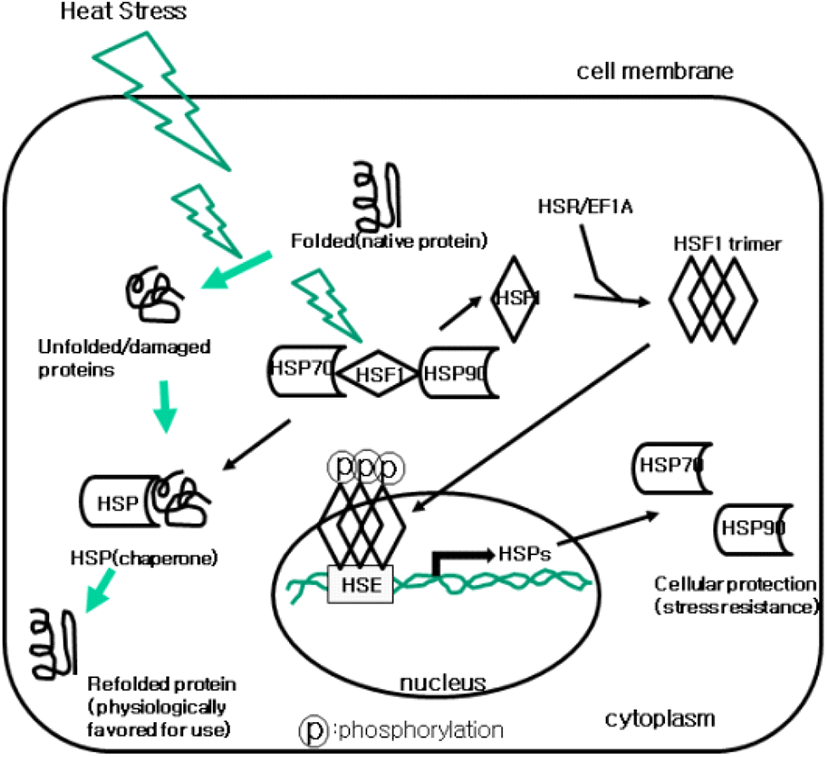
열 내성(thermal tolerance)은 주변 환경의 온도가 체온의 온도중성영역(thermoneutral zone; TNZ)을 벗어날 때 열의 생성과 방출의 균형을 잡는 동물의 능력이다(Hassan et al., 2019). 동물이 몸의 열생산과 손실간의 균형을 유지하기 어려울 때 항상성이 무너지면서 열 스트레스가 발생한다(Greene et al., 2019). 따라서 TNZ는 최소한의 대사적 조절에 의해 몸의 체내온도를 유지할 수 있는 외부환경의 온도범위가 된다. 외부 환경 온도가 이 TNZ 영역에 있는 동물은 정상적 체온 유지를 위하여 요구되는 에너지 소비가 거의 없기 때문에 가축은 최대한 TNZ영역 내에서 사육될 수 있도록 하여야 한다. 포유동물과 조류는 체온을 유지하는 잠재적 능력을 가지고 있으며, 세포수준에서 항상성(homeostasis)을 유지하기 위해서는 특화된 HSP가 있어야 한다. 이러한 단백질들 중에서 HSP70이 가장 중요하다. HSP 단백질들은 열대 혹은 하절기 고온 기후지역에 있는 동물이 열 스트레스에 의한 급격한 변화에 대처하는 방어 역할을 한다. 만약 열 스트레스에 대한 이러한 방어기작이 없다면 상승한 주변 환경온도에 노출된 동물은 치명적 희생자가 될 수 있다(Mishra and Palai, 2014). HSP70은 열 스트레스 하에서 폴리펩타이드의 변성을 막기 위한 중요한 세포의 온도저항성 단백질이다. 만약에 세포가 스트레스에 직면하게 되면 세포내에 물질 수송뿐만 아니라, 단백질의 잘못된 접힘 등에 의하여 세포는 정상적 활동을 못하게 된다. 스트레스 하에서는 HSP가 활성화되어지고, 세포내에서 변성과 비정상적 단백질의 축적을 최소화하여 세포를 구제하는 기능을 하게 된다. 이러한 HSP의 기능은 세포의 생존율을 높이고, 열 스트레스를 극복하는 능력을 제공해 준다(Mishra and Palai, 2014).
동물은 생리적 조화 즉, 생존과 항상성 유지를 위하여 열 스트레스에 대한 적응 혹은 완화를 위한 방어기작이 필요하다. 소, 양, 염소, 닭 등을 포함한 대부분의 동물들은 열 스트레스를 받게 되면 HSP70의 발현이 증가하게 된다(Yu et al., 2008; Mishra Romero et al., 2013; Dangi et al., 2014; Palai, 2014). 열 스트레스가 세포내 HSP의 발현을 유도하고, HSP는 열 스트레스의 부정적 영향을 완화시키기 위한 첫 번째 방어선이 된다(Trinklein et al., 2004; Page et al., 2006). 이러한 HSP의 결정적 역할 때문에 HSP를 열 내성 및 기후변화 저항성과 연계시키게 되고, 가축은 보다 더 열악한 환경적 열 스트레스에서도 적응을 하게 된다(Feder and Hofmann, 1999).
열 스트레스로부터 세포를 보호하기 위해서는 HSP70 단백질이 열 스트레스를 받은 단백질과 결합을 하여 세포내에서 응집이 일어나지 못하도록 하는 것이다. 열 스트레스에 의한 변성된 단백질들 간의 응집은 세포내에서 여러 복잡한 일들을 유도하여 심각한 결과를 초래하게 된다. 따라서 열 저항성은 세포내 HSP의 발현과 이에 의해 생성된 HSP단백질의 총량과 직접적으로 연관이 있어 HSP가 많이 만들어지면 열 저항성이 높다. 반대로, HSP단백질의 분해가 높게 일어나면 열 저항성은 감소하게 된다. HSP70은 스트레스에 의해 유도되어 발현될 수 있으며, 이렇게 생성된 HSP단백질에 의해여 생물은 보다 더 열 저항성을 가지게 된다(Browder et al., 1998). HSP의 발현에 중요한 역할을 하는 전사인자 heat shock factor(HSF)는 평상시 세포내에서 불활성 상태로 존재하는데, 열충격을 받게 되면 번역후 변형과정을 통하여 활성형으로 변하며, 활성형 HSF는 HSP의 전사를 증가시킨다(Sorger et al., 1987). 열 스트레스에 의한 HSP를 생성하는 세부적 과정을 살펴보면, 열 충격에 의해 변성된 단백질은 HSF와 결합되어 있는 HSP를 세포질 내에서 분리시킴으로써 HSF를 활성화시킨다. HSP로부터 분리된 활성형 HSF는 인산화(phosphorylation)가 되어 삼합체(trimer)를 형성한다. 인산화된 HSF삼합체는 핵 안으로 이동하여 열충격단백질유전자(hsp genes)의 프로모터 상에 있는 열충격요소(heat shock elements)와 결합한다. 이의 결과로 hsp 유전자들은 전사(transcription)가 되고, HSP 단백질들을 합성하게 된다(Sorger et al., 1987; Kiang and Tsokos, 1988) (Fig. 1). 지금까지 알려진 HSF는 4종류의 단백질 동형(isoforms), 즉 HSF1, HSF2, HSF3, HSF4가 있으며, 사람에는 HSF1, HSF2, HSF4가 확인되었다(Snoeckx et al., 2001). 닭에서는 HSF1과 HSF3 2종류가 확인되었으며, 현재까지 알려진 HSF3는 오직 닭에서만 보고되었다(Archana et al., 2017). 이들 4종의 HSF들은 세포내 다양한 종류의 스트레스에 의해 활성화 되지만, HSF1의 경우 특히 온도변화에 매우 민감하게 반응하고 활성화 되어 HSP70 단백질의 생성에 주요 역할을 한다(Cotto et al., 1996). 닭에서 HSF1은 중간 열 스트레스에서 그리고 HSF3는 만성 열스트레스에서 활성화가 많이 된다(Pardue et al., 1992; Tanabe et al., 1997). 또한 닭의 HSF1은 오직 HSP70만 유도하는 반면(Inouye et al., 2003), HSF3은 모든 HSP들의 발현을 촉진한다(Tanabe et al., 1998). 합성된 HSP들은 세포질에서 ATP의 도움을 받아 C—말단부위의 HSP와 변성된 단백질과 결합하여 기능적으로 회복된 3차원적 구조 단백질을 갖도록 도와주어 구조적, 기능적 보존 역할을 한다.
열 스트레스에서 HSP70의 주요 기능은 치명적 열 스트레스로부터 세포의 사멸을 억제함에 있다. 가축에서 열 스트레스의 상태에 놓이게 되었을 때 세포는 생존을 위한 반응 체계를 작동한다. 세포 생존 반응의 최종 산물중 하나가 HSP70과 같은 HSP의 합성과 방출이며, 이들은 동물이 열 스트레스로부터 벗어나 생존할 수 있도록 한다. 열 스트레스를 받게 되었을 때, 세포가 스트레스로 인한 부정적 영향을 억제하기 위하여 HSP70의 발현이 높게 나타난다는 증거들은 여러 축종에서 보고되었다(Deb et al., 2013; Romero et al., 2013; Dangi et al., 2014; Kishore et al., 2014). 각각 다른 동물에서 HSP70의 발현의 차이는 종간에 열 저항성의 차이로 볼 수 있다(Agnew and Colditz, 2008). 사람의 평균 체온은 37°C인데 반하여 면양은 39.5°C, 소는 38.9°C로서, 각 동물의 체온의 차이는 최적의 열충격 단백질의 발현에도 영향을 미칠 수 있을 가능성이 있다. 즉, 사람의 백혈구에서 HSP70의 최적 발현은 42°C인데 반하여(Dressel and Gunther, 1999), 소와 면양은 43.5°C에서 최적 발현을 보였으며, 같은 조건에서 면양은 소보다 HSP70의 발현이 더 높았다고 한다(Agnew and Colditz, 2008). 이러한 관점에서 보면 HSP70은 열 내성 가축 선발육종을 위한 중요한 유전적 마커가 될 수 있다. 기후변화가 진행되는 상황에서 고려해 보면, 가축의 열 저항성을 향상시키기 위한 것으로 즉, 전반적인 기후 온난화와 연관한 부정적 영향을 완화시키고, 기후변화에 더 잘 견딜 수 있는 가축의 선발 육종이 필요하다. 이러한 의미에서 세포의 열 저항성과 hsp 유전적 변이의 연구가 주목을 받고 있다. 일반적으로 HSP70은 각기 다른 스트레스 자극에 대하여 유도되어지고 모든 동물에 존재하지만, 각 동물들은 열 스트레스에 대하여 다양한 발현의 차이를 나타낸다. 이는 hsp70 유전자의 5’-또는 3’-UTR flanking 영역에서 자연적으로 야기된 염기의 변이가 발현 차이를 일으키는 이유 중 하나가 될 수가 있다. 이러한 유전적 변이는 스트레스 자극에 대한 유도 능력에 대한 차이, 그리고 hsp70 mRNA의 발현과 안전정의 정도에 영향을 준다. 이러한 결과는 각 동물의 열 스트레스에 대한 저항성의 크기에도 영향을 줄 수 있다(Archana et al., 2017). 소의 HSP70 프로모터 영역에서의 SNP와 열 저항성(Flynn et al., 1991; Deb et al., 2014) 돼지의 HSP70의 3’-UTR 영역의 polymorphism과 연관된 mRNA의 안전정 제고와 열저항성(Schwerin et al., 2002) 등에서 열 저항성이 있는 가축 선발을 위한 HSP70의 활용 가능성을 보여 주었다. 닭에서 열 저항성 관련 hsp70유전자의 SNP에 대한 유전적 효과를 보면, 급성 열 스트레스를 받은 닭의 심장과 간 조직에서 hsp70 유전자의 5’-flanking 영역의 —69A>G 지역의 SNP 즉, GG 유전자형이 AA형보다 더 높은 hsp70 유전자 발현을 보였다(Chen et al., 2016). 이는 hsp70유전자의 프로모터 영역의 SNP가 열 저항성 개체를 선발하기 위한 분자적 마커로서 활용 가능성을 보여준다. 위와 같은 연구들은 동물들 품종들 간에 유전적 혹은 표현적 다양성과 열 저항성과의 관계를 볼 때 매우 의미 있는 결과임을 알 수 있다. 다음 세대를 위한 유전적 개량 즉 열 저항성을 가진 개체의 선발 이용은 가축 생산을 높이기 위한 관리적 측면보다 지속 가능한 가축 생산을 위해 중요한 전략이 될 수 있다. 따라서 HSP70유전자의 UTR flanking영역에 있는 다양한 변이에 대한 연구는 기후 변화에 보다 더 잘 적응하는 개체의 선발을 위한 유전적 마커로 활용되어 동물의 전체적 능력 향상뿐만 아니라, 동물 복지 측면까지 향상 시키는데 결정적 도움을 줄 수 있다.
동물의 다양한 세포에서 열 스트레스에 맞서기 위해 HSP70을 발현한다. HSP70은 지속적 발현과 유도성 발현으로 구분할 수 있다. 유도성 HSP70발현은 세포질에서 분자 샤프론의 활성으로 스트레스를 완화시키는 기능을 한다(Nollen et al., 1999). 유도성 HSP70은 가축의 스트레스저항 능력과 연관이 높기 때문에 동물의 혈청에서 열 저항성에 대한 바이오마커로서 HSP70 단백질의 수준을 측정하면 세포의 열 스트레스에 대한 정도를 판단할 수 있다(Flanagan et al., 1995). 소, 염소, 양, 닭 등의 열 스트레스를 받은 가축에서 유도형 HSP70의 발현이 증가된 것을 확인하였다(Yu et al., 2008; Zulkifli et al., 2010; Mishra et al., 2011; Dangi et al., 2012). 가금에서도 열 스트레스의 반응의 결과를 확인하기 위하여 다양한 평가방법들이 소개되었다. 열 스트레스를 완화시키기 위한 다양한 전략들을 찾기 위하여 가금에서 많이 적용하고 있는 스트레스 평가 방법은 corticosterone과 같은 스트레스 호르몬과 heterophils과 lymphocytes의 비율(H/L ratio) 그리고 텔로미어의 마모 및 DNA 손상 정도(Sohn et al., 2012; Bateson, 2015) 등을 주요 표준 바이오마커로 활용하고 있다. 일반적으로 닭의 corticosterone의 혈중 농도는 낮은 수준(0.3∼20 ng/mL)으로 있지만(Zhang et al., 2009; Kang and Kuenzel, 2014), 스트레스를 받은 닭의 혈중 농도는 최고 150 ng/mL(Scanes, 2016)까지 높다. 스트레스 호르몬의 혈중 농도는 닭의 생리적 상태(번식주기/산란주기), 시료 채취시간 및 개체별 차이 등에 따라 많은 변이를 보여 스트레스 수준을 해석하는데 영향을 줄 수 있다. 또한 혈액 채취를 위한 닭의 고정하는 그 자체만으로도 스트레스를 유발할 수 있다. Heterophils과 lymphocytes의 비율은 질병과 감염에 의한 영향을 많이 받으며, 외부 환경으로부터 오는 스트레스에 대한 진정한 스트레스의 수준을 반영하기에는 미흡한 부분이 있다(O’Dell et al., 2014). 텔로미어의 분석을 통한 조류의 스트레스 평가방법은 조직 혹은 유핵적혈구(nucleated erythrocytes) 등을 이용할 수 있지만, 대체적으로 lymphocytes를 분석하여 텔로미어의 마모 정도 혹은 DNA손상 등을 비교한다(Sohn et al., 2012). DNA손상은 급성 스트레스를 평가하는데 비교적 적합하고, 텔로미어를 이용한 스트레스 분석은 만성 스트레스 또는 일생동안 축적된 스트레스를 평가하는데 유용하다고 할 수 있으며, 특히 동물의 복지를 중요시한 장기적 사양체계를 평가하는데 적합하다(Sohn et al., 2012; Bateson, 2015). 최근에는 비-침습성(non-invasive)이면서 재현성과 정확성이 높은 깃털 HSP70을 분자수준에서 측정하는 방법이 제시되었다. 브로일러의 경우, 다른 주령보다 6∼8주령 사이에 가장 열 스트레스에 민감한 것으로 알려져 있다(Temim et al., 2000). Greene 등(2019)은 6주령의 육계에서 여름 사육기간의 아침(24°C)과 오후(29.5°C)에 성장하는 깃털(growing feather)을 취하여 HSP70의 mRNA 및 단백질 발현을 조사하였다. 오후에 채취한 깃털에서 발현되는 HSP70의 mRNA 및 단백질 모두 아침에 취한 깃털보다 이 유전자의 발현이 모두 높게 발현하였다. HSP70 단백질의 발현을 소장 및 혈액에서 분석하였을 때에도 깃털에서 발현된 양상과 같은 패턴을 보여 줌으로써, 성장하는 깃털을 이용한 HSP70의 발현 확인 방법이 비-침습성이면서 닭의 열 스트레스 정도를 세포수준 혹은 동물 개체수준의 스트레스를 반영할 수 있고, 반복적, 체계적으로 분석할 수 있는 분자적 마커로 활용할 수 있음을 보여주었다. HSP70은 체내 장기와 혈액, 깃털 등에 전반적으로 발현되고, 갈증, 감염, 운송, 추위 등과 같은 다양한 스트레스 인자들에 의해 발현되기 때문에 깃털 HSP70의 분석은 가금류의 스트레스 수준을 체계적으로 평가 하는데 이용될 수 있다. 성장 중인 깃털을 이용한 HSP70의 분석은 깃털의 펄프(pulp)부위를 이용하게 된다(Fig. 2). 조류의 성장 중인 깃털의 펄프(pulp) 부위는 약 10 mm 정도의 깃털 하단 부위로 표피층(epidermal layer)과 결합조직(connective tissue)에 의해 발달된 내 진피(inner dermis)로 구성되어 있다. 이 펄프에는 동맥혈관과 모세혈관이 잘 형성되어 있으며, 백혈구, 면역세포, 림프계 및 신경세포, 각종 구조세포 등으로 이루어져 있다(Yu et al., 2004). 깃털 여포(feather follicle)가 진피유두세포 조직(dermal papilla)으로 모래시계 모양을 하고 있다. 진피유두세포조직을 싸고 있는 부위를 칼라(collar)라고 하며, 이 부위의 밑부분이 세포분열 영역이 된다. 칼라부위가 세포분열을 하여 깃털 필라민트의 각질 세포가 된다. 따라서 가금 깃털의 펄프는 가금에게 스트레스를 최소화 시키면서 연속 혹은 반복적 실험을 위한 주요한 시료 확보 장소가 될 수 있다.

결 론
가축에서 고온 환경은 생리적 측면뿐만 아니라, 생산능력에도 해로운 영향을 준다. 동물은 해로운 영향을 억제하고 스트레스로부터 자신을 보호하기 위해 여러 가지 기작들을 작동하고, 그 중심에는 HSP70이 있다. 이러한 기작을 세포수준에서 잘 이해하고 스트레스를 체계적으로 평가한다면 급격한 기후변화 시대에 잘 적응할 수 있는 가축을 선발하고 지속 가능한 축산업으로 발전할 수 있으며, 이는 동물복지에도 기여할 수 있다. 가축이 짧은 시간 혹은 만성적 열 스트레스에 노출되었을 때, 스트레스 조건에 따른 쉽고 신뢰성 있는 스트레스 평가 방법의 확립이 필요하다. 또한 가축별 품종별 열 스트레스 연관 후성 유전체 분석을 체계적으로 진행한다면 열 저항성 가축을 선발하는데 크게 기여할 것이다.