
서 론
대식세포(macrophage)는 내피와 조직구(histocytes)에 있으며, 주로 식균 작용에 관여한다(Van Furth et al., 1972). 단핵식세포계(mononuclear phagocyte system, MPS)는 수상돌기세포(dendritic cells)와 후에 혈액단핵세포로 분화하여 대식세포가 되는 골수의 전구세포로 정의할 수 있다(Hume et al., 2002). 대식세포는 척추동물의 모든 조직에 염증을 촉진하는 곳에서 중요한 기능을 갖는다(Geissmann et al., 2010). 대식세포의 발달은 대식세포 집락 자극 인자(M-CSF, colony stimulating factor 1(CSF1))가 필요한데, 골수 전구 세포에 있는 MCSF 수용체(M-CSFR, 또는 CSF1R)에 대한 결합을 통해 이루어진다(Mouchemore and Pixley, 2012). 대식세포는 면역 및 상처 치유 기능을 통해 거의 모든 질병, 특히 암 및 비만과 같은 만성 질환에서 중요한 조절기능을 갖는다(Pollard, 2009). 최근의 CSF에 대한 연구는 치료의 특정 표적화를 위한 기회를 줄 수 있으며(Hamilton, 2008), M-CSF의 효과는 세포 표면 수용체인 M-CSFR에 의해 매개되며, homodimer 또는 heterodimers로서 리간드-수용체 결합을 통하여 신호 전달 조절 역할을 한다(Chen et al., 2008).
최근에 IL-34는 포유류, 조류, 그리고 어류에서 M-CSFR에 대한 두 번째 기능적 리간드로 확인되었다(Haishan Lin, 2008; Garceau Valerie et al., 2010; Wei et al., 2010; Wang and Colonna, 2014). M-CSFR 유전자는 대식세포에서 발현되며, IL-34 및 M-CSF 리간드는 대식세포 증식을 촉진할 수 있다. 수용체와 두 리간드의 공진화(co-evolution)에 대한 분석은 수용체와 리간드가 서로 다른 위치에서 독립적으로 결합할 수 있음을 나타낸다(Garceau Valerie et al., 2010).
인터루킨(IL)은 면역 체계에서 중요한 역할을 하는 사이토카인 그룹으로, 림프구 및 기타 세포의 성장, 이동성 및 분화를 조절하여 염증과 면역을 조절한다. IL-34는 특별히 비장 조직에서 발현되며, 인간 단핵구 생존을 지원하고 대식세포 형성을 촉진한다(Chemel M et al., 2012; Hwang et al., 2012). IL-34(IL34LacZ/LacZ mice)는 접촉 과민증을 촉진하는 랑게르한스 세포와 바이러스 감염과 관련된 손상으로부터 중추 신경계를 보호하는 미세아교세포(microglia)에서 중요한 역할을 한다고 보고되었다(Wang and Colonna, 2014). IL-34는 분비된 단백질 라이브러리의 기능적 스크리닝을 통해 확인되었으며(Haishan Lin, 2008), 이 사이토카인은 M-CSFR을 통해 대식세포의 분화와 생존을 촉진한다. 그리고 M-CSFR에 대한 결합을 통해 IL-34는 M-CSF와 유사한 기능을 공유한다. 또한 이 두 사이토카인은 사람의 혈액내 케모카인의 발현을 유도하고(Eda et al., 2010), 대식세포의 항상성을 유지해주는 역할도 한다(Munoz-Garcia et al., 2021). M-CSF 프로모터의 제어 하에 IL-34는 뼈 및 조직의 대식세포를 복구할 수 있다(Wei et al., 2010).
IL-34 mRNA는 심장, 뇌, 폐, 간, 신장, 비장, 흉선, 고환, 난소, 소장, 전립선, 결장을 포함한 성인 인간 조직에서 광범위하게 발현된다(Haishan Lin, 2008). IL-34와 M-CSF의 발현은 IL-1β 및 TNF-α 신호전달과 같은 전염증성(proinflammatory) 사이토카인에 의해 유도되며, M-CSF는 최근 닭(Garceau et al., 2015; Garcia-Morales et al., 2014)과 금화조(zebra finch)(Garceau Valerie et al., 2010)에서 발견되었다.
본 연구에서 포유류와 조류의 IL-34에 대한 유사성을 분석하고, 닭의 다양한 병원균 감염 조직 및 세포주에서 IL-34의 발현을 비교 분석하였다. 또한 시험관 실험(in vitro)에서 M-CSFR 리간드인 M-CSF와 함께 IL-34의 발현을 관찰하였으며, 두 개의 리간드가 LPS에 의해 차등적으로 발현되고 조절된다는 것을 보고하였다. 그리고 E. coli로부터 정제된 재조합 닭 IL-34 및 M-CSF는 닭 대식세포 세포주에서 M-CSFR의 발현을 촉진하는 강력한 생물학적 활성을 관찰하였다.
재료 및 방법
중앙대학교 사육실에서 5주령 ROSS 308 육계 수컷(양지 부화장, 평택) 중 독립적으로 설 수 없는 관절 이상 개체 4마리와 정상적인 개체 4마리에서 흉선, 비장, 소장, 맹장 및 총 배설강 등의 조직에서 RNA을 추출하여 사용하였다. ADOL 6.3, ADOL 7.2 line(Avian Disease and Oncology Laboratory, USDA-ARS), 그리고 White Leghorn 닭의 각 조직 total RNA을 미국 농무성 ARS의 ABBL(Animal Biosciences and Biotechnology Laboratory, Beltsville, MD)에서 공여 받았다. 2개의 근친 계통인 ADOL 6.3(resistant)과 7.2(susceptible) 계통은 avian leucosis virus과 Marek’s disease virus에 저항성 및 감수성 계통으로 육종되었으며(Bacon et al., 2000), 이 계통을 이용하여 21일령에 E. maxima strains(103 to 104 oocysts/bird) 감염 후, 5일 후에 C. perfringens(1 × 109 cfu/bird)을 추가 감염시켜 괴사성 장염(necrotic enteritis, NE)을 유도하여 비장(spleen) 및 소장(small intestine) 조직을 추출하였다(Lee et al., 2011). 6개월령의 White Leghorn에 S. Enteritidis(1 × 108 CFU/bird) 감염 7일 후에, 폐(lung), 난소(ovary), 소장, 비장, 간(liver), 흉선(thymus), 그리고 난관(oviduct) 조직을 추출하였다. 모든 실험은 Beltsville Agricultural Research Center의 Institutional Animal Care and Use Committees의 승인 하에 수행되었다. 1일령 ROSS 육계는 무항생제/무항콕시듐제의 사료와 물을 무제한 급여하였으며, 실험 동안 일정한 광원 하에 관리되었다. E. coli O111:B4 유래의 Lipopolysaccharides(LPS, Sigma-Aldrich, St. Louis, MO)는 phosphate-buffered saline (PBS, equivalent to 1 mg/ml)에 희석하여 사용하였다. 2주령의 각 6수 암컷 및 수컷 육계에 0.2 mg/BW/kg의 LPS을 피하 주사하였으며, 대조구는 PBS을 주사하였다. 1일 후 총 20수에서(처리구 암/수 5마리, 대조구 암/수 5마리) 심장, 간, 비장, 신장, 흉선, 췌장(pancreas), 십이지장(duodenum), 공장(jejunum), 회장(ileum), 맹장(caeca), 총배설강(cloaca), 쓸개(gall bladder), 후두(larynx), 기도(trachea), 그리고 폐 조직 등을 추출하였다. 이 실험은 중앙대학교 동물실험위원회(IACUC) 승인(No. 2016-00108)하에 수행되었다.
닭의 IL-34와 M-CSF 프라이머는 predicted chicken IL-34 cDNA 염기서열(accession number XM_003641892.3)과 닭 M-CSF cDNA 염기서열(accession number NM_001193295)을 NCBI/primer-blast(https://www.ncbi.nlm.nih.gov/tools/primer-blast/)을 이용하여 디자인하였다(Table 1). 닭의 각 조직을 섞은 cDNA을 Dream Taq green PCR Master mix(Thermo scientific, Waltham, MA)을 이용하여 다음의 조건에서 PCR 증폭하였다; 초기 95°C/5min 반응 후, denaturation 95°C/30s, annealing temperature/30s, 그리고 extension 72°C/1min으로 35 cycles을 수행하였으며, 마지막 72°C/5min 으로 마무리 하였다. PCR 결과물은 PureLinkTM quick gel Extraction Kit(Invitrogen, Carlsbad, CA)을 이용하여 정제 후, pCR2.1-TOPO vector(invitrogen)에 ligation 및 E. coli TOP10 competent cells(invitrogen)에 형질전환하였다. Blue-White 스크리닝을 통하여 positive clones을 선택하여 plasmid 추출 후 염기서열에 대한 sequencing을 실시하여 확인하였다(Genotech, Daejeon, Korea).
닭 IL-34/pCR2.1-TOPO와 닭 M-CSF/pCR2.1-TOPO을 제한효소 EcoRI와 HindIII(Promega Corporation, Fitchburg, WI)으로 처리 후, PureLinkTM quick gel Extraction Kit (invitrogen)으로 정제하였다. 재조합 단백질 발현 벡터인 pET32a E. coli expression vector(Novagen, Cambridge, MA)에 ligation 후 E. coli BL21(invitrogen)에 형질전환하였다. 염기서열 확인 후, 박테리아를 ampicillin을 포함한 LB media (Thermo scientific) 20°C 온도에서 0.5 mM isopropyl β-d-1-thiogalactopyranoside(IPTG, USB Cleveland, OH)을 처리하여 overnight(250 rpm) 배양하였다. 박테리아는 4°C 온도에서 5,000 × g로 20분간 원심분리하여 모은 후, B-PER Bacterial Protein Extraction(Thermo Scientific)을 이용하여 처리하였다. 재조합 단백질 IL-34와 M-CSF는 HisPurTM Ni-NTA Resin(Thermo Scientific)을 이용하여 정제하였으며, 3000-molecular weight cut-off membrane(Amicon Ultra-15 Centrifugal Filter Unit; EMD Millipore, Temecula, CA)을 이용하여 농축하였다. 재조합단백질은 SnakeSkin Dialysis Tubing(Thermo Scientific)을 이용하여 PBS(pH 7.0)에서 정제하였다.
재조합 단백질은 sample buffer(0.125 M Tris-HCl [pH 6.8], 4% SDS, 20% glycerol, 10% 2-mercatoethanol, and 0.004% bromophenol blue)와 같은 비율로 섞은 후, 95°C에서 5분간 열을 가하였으며, 10% SDS-polyacrylamide gel에서 전기 영동 후, PVDF western blotting membrane(Roche, Indianapolis, IN)에 blotting 하였다. Membrane은 5% skim milk를 포함하는 PBS을 이용하여 4°C에서 16시간 동안 blocking 하였다. 0.05% Tween 20(PBS-T)을 포함한 PBS을 이용하여 membrane을 3회 세척 후, Anti-6X His tag® antibody(Abcam, Cambridge, UK)을 이용하여 4oC에서 overnight 배양하였다. Membrane은 PBS-T을 이용하여 5회 세척 후, Pierce ECL western blotting substrate(Thermo Scientific) 및 Amersham hyperfilm ECL high performance chemiluminescence film(GE Healthcare, Buckinghamshire, UK)과 8 × 10” X-ray cassette(Soyee product inc, Republic of Korea)을 이용하여 확인하였다.
Expert Protein Analysis System(ExPASy [www.expasy.org/tools/])을 이용하여 단백질에 대한 분자량과 이론적 등전점(theoretical isoelectric point, pI)을 확인하였다(Gasteiger et al., 2005). 또한 signal peptide sequence는 SignalP 4.1 software program(http://www.cbs.dtu.dk/services/ SignalP) 을 이용하여 예측하였다(Petersen et al., 2011). CLUSTAL Omega software(http://www.ebi.ac.uk/Tools/msa/ clustalo/)을 이용하여 아미노산 다중정렬(multiple alignments)을 수행하였으며, disulfide bond를 DiANNA 1.1 software(http://cla vius.bc.edu/~clotelab/DiANNA/)로 예측하였다(Ferrè and Clote, 2005). 유사성 분석(homology analysis)은 MatGat software(http://ww3.bergen.edu/faculty/jsmalley/matgat.html)을 이용하였으며(Campanella et al., 2003), 닭 IL-34에 대한 단백질 구조 분석(PDB)은 I-TASSER(http://zzanglab.ccmb.med. umich. edu/I-TASSER)을 이용하여 mouse IL-34(PDB Code c4exnF)을 이용하였으며, 그림은 Genome3D의 Phyre2 (http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index)(ProteinHomology/analogY Recognition Engine V 2.0)을 이용하여 수행하였다(Kelley et al., 2015).
닭 대식세포주인 HD11는 10% fetal bovine serum, 10,000 U/mL penicillin streptomycin(HyClone, Waltham, MA)이 포함된 RPMI 1640(Gibco, Carlsbad, CA) 완전배지에서 배양하였다. 모든 세포는 4 × 106 세포를 41°C 온도와 5% CO2 조건하에서 60 mm culture dish(Corning)에서 배양하였다. HD11세포는 1 μg/mL lipopolysaccharide(LPS from E. coli O111:B4; Sigma-Aldrich)을 이용하여 0, 2, 4, 6, 8, 12, 그리고 24 h 동안 처리 후 TRIzol(Ambion, St’Austin, TX) 처리하여 total RNA을 추출하였다.
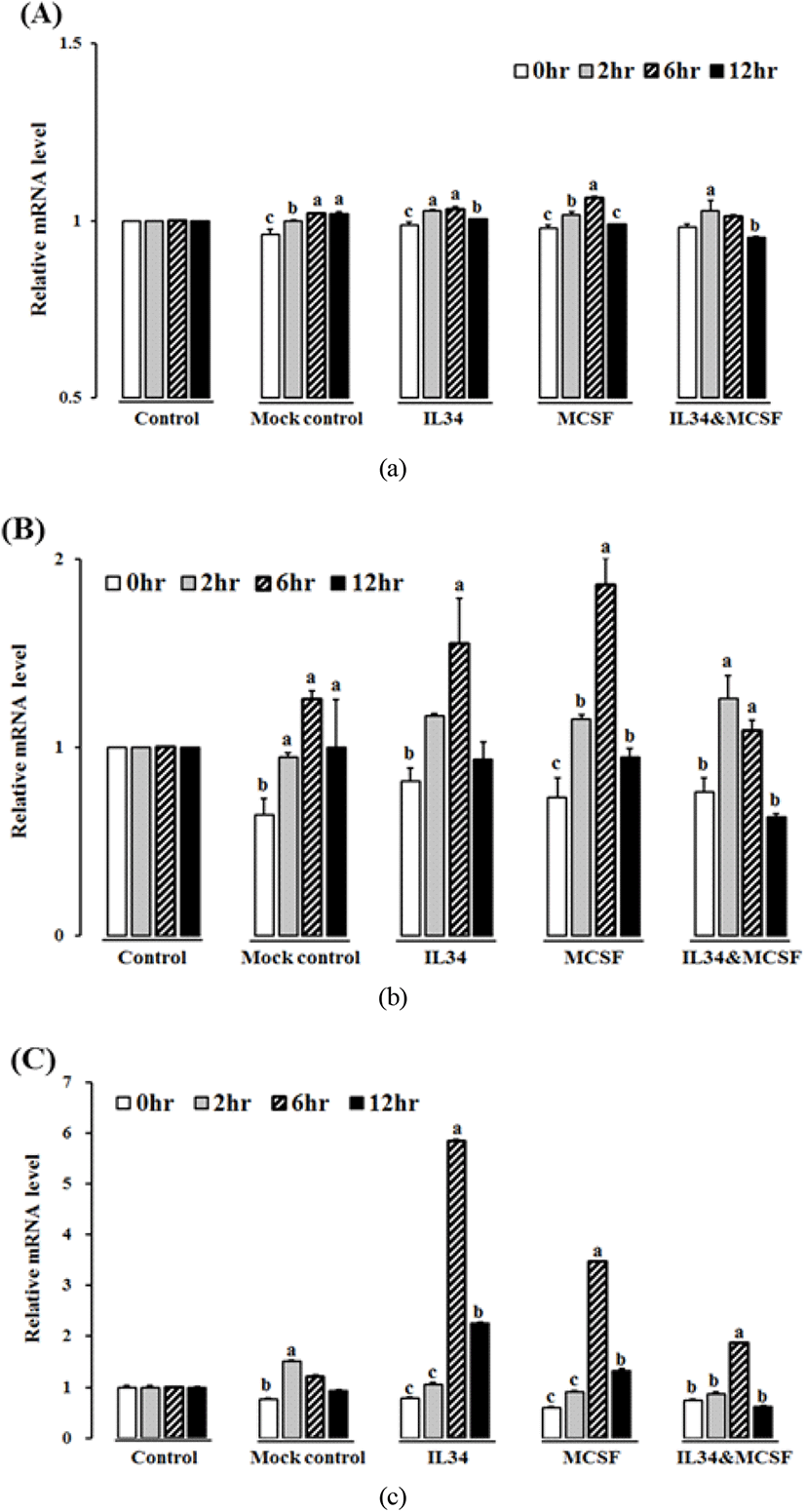
닭 IL-34와 M-CSF는 in vitro에서 표적 세포를 발현하는 M-CSFR에 대해 유사한 효과를 나타내므로 세포 내에서도 유사한 발현 패턴을 보이는지를 조사하였다. 재조합 단백질인 닭 IL-34(200 ng/mL), 닭 M-CSF(200 ng/mL), 두 단백질 IL-34(100 ng/mL)와 M-CSF(100 ng/mL), medium, 그리고 mock control(TRX; pET32a vector, 100 ng/mL)을 대조구로 사용하였다.
각 재조합 단백질의 최적 농도는 사전 실험을 통하여 정하였다(unpublished data). 각각의 처리구는 처리 후 0, 2, 6, 12, 24, 그리고 48 h 후에 TRIzol(Ambion)에 녹여 total RNA을 추출하여 quantitative real-time PCR(qRT-PCR)을 이용한 발현 분석에 사용하였다.
HD11 macrophage 세포주와 ADOL 6.3/7.2 라인의 조직(thymus, spleen, intestine, caeca, cloaca), White Leghorn(lung, ovary, small intestine, spleen, liver, thymus, oviduct) 및 ROSS 308 육계의 각 조직(heart, liver, spleen, kidney, thymus, pancreas, duodenum, jejunum, ileum, caeca, cloaca, gall bladder, larynx, trachea, lung)으로부터 TRizol을 이용하여 RNA을 추출하였다. cDNA합성을 위하여, 각 total RNA 2 μg을 DNase I(Thermo Scientific)을 처리 후, RevertAid First Strand cDNA Synthesis Kit(Thermo Fisher Scientific)을 이용하여 cDNA을 합성하여 유전자 발현분석에 사용하였다.
합성된 cDNA는 nuclease-free water을 이용하여 100 ng/μL의 농도로 희석하였으며, 100 ng template cDNA, 2 × qPCR mix Lo-ROX(ENZO Life Science, Farmingdale, NY), 5 pmole forward/reverse primers(Table 2)로 반응액을 조성 후 LightCycler 96(Roche)을 이용하여 다음의 반응조건하에서 qRT-PCR을 수행하였다. Pre-heating 95°C/10 min, denaturation 95°C/10 sec, annealing 52~58°C/10 sec, extension 72°C/15 sec으로 45 cycles로 수행하였다(Table 2). 유전자 발현에 대한 상대적 값은 GAPDH 유전자를 이용하여 2−ΔΔCt 방법으로 계산하였다(Thomsen et al., 2010).
관절이상과 정상개체간의 조직별 비교(Fig. 1)와 산란계와 육계의 조직별 처리에 대한 IL-34발현 비교(Fig. 2)는 SPSS을 이용한 Student t 검정, LPS처리에 대한 유전자의 시간대별 비교 및 재조합 단백질 처리에 대한 세포주의 시간대별 유전자 발현비교(Fig. 3, 6)는 일원분산분석(ANOVA)으로 처리 간 유의성을 검정하고, 처리 평균값 간의 다중검정은 Duncan’s multiple comparison test 검정 방법을 이용하였다. 모든 실험은 최소 3반복 실험을 수행하였으며, 데이터는 mean ± SEM로 표기하였고, 유의수준은 P<0.05에서 유의하게 차이가 있다고 판단하였다.
결 과
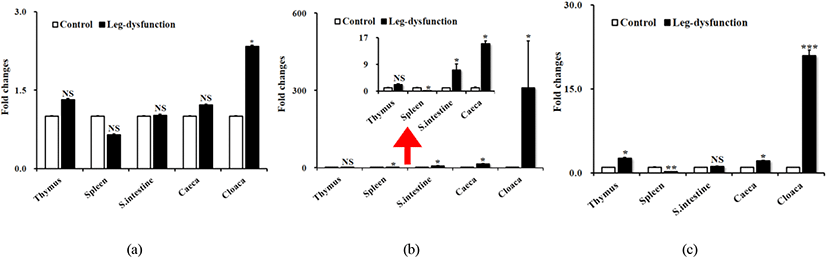
총 5개 조직(thymus, spleen, small intestine, caeca, cloaca)에 대하여 정상인 개체와 관절 이상을 보이는 개체에 대하여 IL-34, M-CSF, 그리고 M-CSFR 에 대한 유전자 발현 분석을 실시하였다(Fig. 1). M-CSF와 M-CSFR 유전자의 경우, 조직간에 많은 발현 차이를 보였으며, 비장에서 가장 낮은 발현을 보인 반면, 총배설강에서 정상 개체와 이상 개체간에 가장 큰 발현 차이를 보였다. IL-34의 경우, 총배설강을 제외한 나머지 조직에서는 유의성을 보이지 않았다. Spleen의 M-CSF와 소장의 M-CSFR 유전자를 제외하고는 거의 대부분의 조직에서 유의적으로 차이가 있음을 보였다.
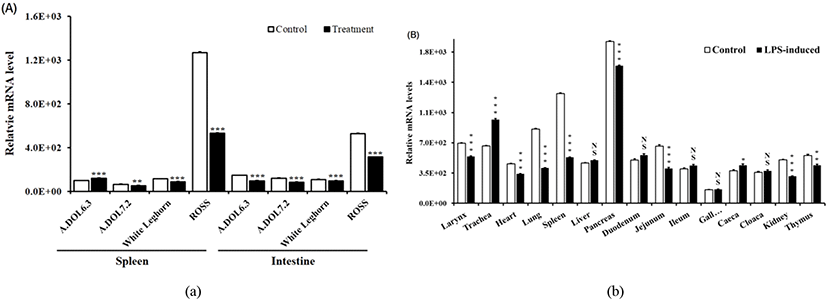
괴사성장염(NE, E. maxima, C. perfringens), S. Enteritidis, LPS 그리고 대조구의 비장 및 소장에서의 IL-34 유전자 발현 비교 결과, ADOL6.3 계통의 비장 조직을 제외하고는 대부분의 조직에서 처리구 대비 대조구에서 유의하게 높게 발현되었다(Fig. 2A). 2주령의 육계 ROSS의 경우, 특히 산란계 계통인 ADOL과 WL에 비하여 전체적으로 높게 발현되었으며, 조직별 비교에서는 8개 조직, larynx, heart, lung, spleen, pancreas, jejunum, kidney, 그리고 thymus에서 처리구보다 대조구에서 높게 발현된 반면, trachea와 caeca는 처리구에서 높게 발현되었다(Fig. 2B).
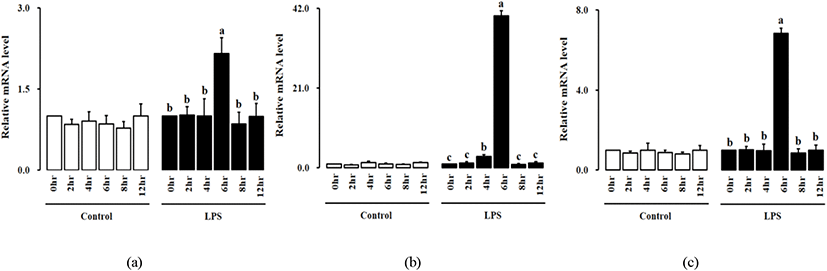
닭의 대식세포인 HD11 세포주에 LPS처리 후, 0, 2, 4, 6, 8, 12, 그리고 24 h 동안 닭 IL-34, M-CSF, 그리고 M-CSFR 유전자의 발현 분석을 실시하였다(Fig. 3). 모든 처리구에서 처리 6시간 후에 IL-34, M-CSF 그리고 M-CSFR mRNA의 발현이 유의적으로 높게 나타났다.
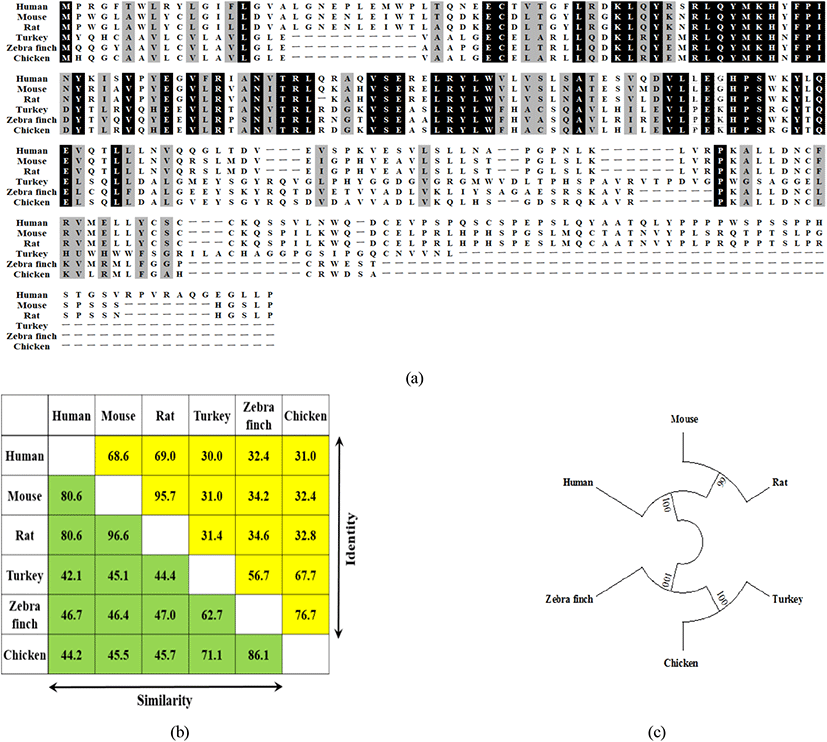
닭의 각 조직을 섞은 cDNA을 이용하여 537bp coding sequence으로 이루어진 178-아미노산, 20.2kDa의 chIL-34을 클로닝하였다. 아미노산 서열은 MatGat software(Campanella et al., 2003)을 이용하여 타 종과 비교 분석결과, 칠면조(67.7%)와 금화조(zebrafinch, 76.7%)에 대하여 높은 유사성을 보였으나, 포유동물과는 31.0-32.8%로 낮은 유사성을 보였다(Fig. 4).
닭 IL-34은 N-terminal 1~23 아미노산의 signal peptide을 가지며, “up-down-up-down-up-down” core 구조를 갖는 짧은 beta strand와 6개의 긴 alpha helices bundle(aA-aF)로 이루어진다(Liu et al., 2012; Ma et al., 2012)(Fig. 5B). 추가로 IL-34는 6개의 six-helix bundle 중 첫번째와 마지막 6번째간의 Cys25-Cys161 disulfide bridge(alpha A and alpha F)가 있으며, Cys5-Cys10 와 Cys88-Cys173간에도 disulfide bridge가 있다. 즉 6개의 cysteine이 3개의 disulfide bridges 1-2, 3-5, 4-6(cysteine 5와 10, 25와 161, 88과 173)을 구성하였다(Fig. 5A).

재조합 단백질 IL-34와 M-CSF을 HD11 세포주에 처리하여 각 시간별로(0, 2, 6, 12 h) IL-34, M-CSFR, IFN-γ, 그리고 IL-12 유전자의 발현을 측정하였으며, 벡터의 TRX-tag 단백질을 mock control로 사용하였다. IL-34 cytokine은 모든 처리구와 시간대별 비교에서 큰 차이를 보이지 않았으나, M-CSFR 유전자는 IL-34, M-CSF, 또는 두 단백질 동시 처리구의 6시간에서 유의적인 차이를 보였다(Figure 6A, B). IFN-γ 역시 IL-34, M-CSF, 그리고 두 단백질 동시 처리구의 6시간에서 유의적으로 높게 발현하였다(Fig. 6C). 하지만 IL-12 cytokine은 M-CSF 처리구의 6시에서만 차이를 보였다(data not shown). 4개의 유전자 M-CSF, M-CSFR, IFN-γ, 그리고 IL-12 모두 12시간 이후에 감소하였으며, 전염증성 사이토카인인 IL-1β, IL-6, 그리고 IL-10에 대하여 분석하였으나, 유의적인 차이는 없었다(data not shown).
고 찰
본 연구는 여러 종류의 병원균에 대한 닭 IL-34의 특성 분석에 대한 내용으로 IL-34는 M-CSFR에 대한 또 다른 잠재적 ligand이다. IL-34는 M-CSFR에 결합하며, M-CSF 역시 결합 시 경쟁관계에 있다. IL-34와 M-CSF는 구조적으로 유사하여 M-CSFR의 같은 결합부위를 갖는다(Lin et al., 2008; Garceau V. et al., 2010). 본 연구에서 포유동물과 조류 IL-34와 비교 시 포유류와는 낮은 유사도(31.0~32.8%), 조류와는 높은 유사도를 보였다(67.7~76.7%). 닭 IL-34 유전자는 6개의 helix bundle이 “up-down-up-down-up-down” folds를 이루는 topological diagram을 보여주었는데, 이는 기존의 mouse CSF-1의 구조를 이루는 four-helix bundle(PDB Code 3ejj)과는 다른 결과이지만(Garceau et al., 2010), mouse IL-34 구조(PDB Code c4exnF)를 기반으로 IL-34의 단백질 구조를 완성하였다.
사람의 관절염(rheumatoid arthritis, RA) 환자의 관절 조직(joint tissue), 혈청(serum), 그리고 관절 낭액(synovial fluid)에서 IL-34 유전자가 많이 발현된다는 연구가 있었으며(Chemel M. et al., 2012; Hwang et al., 2012; Moon et al., 2013; Tian et al., 2013Chang et al., 2015), 관상동맥질환(coronary artery disease) 환자의 혈청(Liu et al., 2012), 비만과 제2형의 당뇨병 환자의 혈청과 지방조직(Chang et al., 2014), 염증성 장질환(inflammatory bowel disease) 환자의 점액, 회장(ileum)과 결장(colon) 조직 등에서도 많이 발현된다(Franze et al., 2015; Zwicker et al., 2015).
닭과 칠면조에서 가장 흔한 화농성 관절염(purulent arthritis)의 경우, E. coli,Staphylococcus, 또는 S. Enteritidis 등에 의하여 발병하며, 패혈증(septicemia)으로 발전한다(Goldenberg, 1998; Oh et al., 2010). 본 연구에서 육안으로 관절 이상 개체를 선별하여 조직별로 분석한 결과, IL-34, M-CSF, 그리고 M-CSFR의 발현이 유의적으로 증가하였지만, 일부 조직의 경우, 오히려 감소한 결과를 보였다. 이는 염증반응이 일어나기 전에 병원균(LPS)이 처리되어 면역관련 유전자가 처리구에서 감소된 것으로 판단된다.
우리는 세포주에서 2개의 리간드(IL-34 and M-CSF)와 하나의 리셉터(M-CSFR)가 LPS 자극 시 다르게 발현됨을 확인하였으며, 관절 이상 닭에서 염증반응의 반응자(responder)임을 확인하였다. 포유류 IL-34 및 M-CSF mRNA의 발현 패턴은 일시적으로 나타나며, 이는 생체 내에서 M-CSFR을 활성화한다(Wang et al., 2012). 닭 IL-34의 발현은 LPS 자극 후, 조직별로 많은 변이를 보였다. 특히, IL-34 또는 M-CSF 단독처리 시 두 단백질 동시 처리보다 높게 나타났다. 닭 IL-34는 in vitro에서 LPS 처리 시 증가하였다.
닭 M-CSFR 유전자는 서로 경쟁하는 IL-34와 M-CSF 단백질에 의해 자극된다(Garceau V. et al., 2010). 닭 M-CSFR 발현 역시 재조합 단백질 IL-34과 M-CSF 단독 처리에 의해 처리 후 6시간 후에 최대치를 보였으나, 병합 단백질 처리는 2시간에 최대치에 도달하여, 아마도 2개의 리간드가 receptor와의 결합(IL-34:M-CSFR, M-CSF:M-CSFR complex)이 영향을 주었을 것으로 판단된다.
대식세포의 면역 기능을 감안할 때 대식세포는 interleukin-12(IL-12)와 같은 전염증성 사이토카인을 방출하여 IFN-γ에 반응한다(Gordon and Martinez, 2010). IFN-γ 발현은 재조합 IL-34와 M-CSF에 의해 증가하였으며, IL-12 역시 M-CSF 단독 처리시 증가하였다. 사람의 경우, M-CSF는 단핵구(monocyte)를 M2 대식세포로 분화시키며, 이 대식세포는 IL-1β, IL-6, TNF-α 같은 전염증성 사이토카인이 아닌 chemokine을 발현한다(Verreck et al., 2006). 또한 M-CSF는 LPS같은 co-stimulator와 더불어 전염증성 사이토카인과 chemokine의 발현을 유도한다(Eda et al., 2010).
대식세포에서 M-CSF에 의한 IP-10, MCP-1, MIP-1β, 그리고 TARC 같은 chemokines의 발현에 대한 보고가 있었으나(Fleetwood et al., 2007; Irvine et al., 2009; Eda et al., 2010), 닭에서는 IL-34와 M-CSF가 IL-1β, IL-6, 그리고 IL-10의 발현을 유도하지 않았다.
결론적으로, 닭의 다양한 품종별, 조직, 세포주, 그리고 관절 이상 닭에 대하여 다양한 질병 병원균을 처리하여 닭의 IL-34에 발현 분석 및 비교 결과, 닭의 다양한 조직 및 세포에서 발현되었으며, M-CSFR의 경우, HD11 세포주에서 재조합 chIL-34와 chM-CSF 단백질에 의해 증가하였다. 이 결과는 다양한 병원균 처리에 대한 IL-34의 발현에 대한 중요한 정보를 제공하며, 추후 닭의 IL-34을 이용한 inflammatory pathway, chemokine 발현 및 체내에서의 역할 등에 대한 연구가 필요하다고 판단된다.