
서 론
초기 병아리의 건강 상태는 이후 개체의 생산능력에 매우 큰 영향을 미친다. 부화 직후 병아리의 강건성은 품종과 같은 유전적 요인에 의해 주로 영향을 받지만 부화 관리적 요인에 의해서도 많은 영향을 받는다. 부화 관리적 요인으로서는 부화 온도나 습도, 전란 등과 같은 부란 관리적 요인과 종계의 산란 연령이나 종란의 보관 상태 등과 같은 부란 외적 요인이 있다. 종계의 품종이나 산란 연령 및 종란의 보관 기간이 부화 능력에 미치는 영향에 대해서는 비교적 잘 알려져 있으나(Choi and Sohn, 2020), 이들 요인들이 발생 병아리의 능력에 미치는 영향에 대해서는 별로 연구된 바가 없다. 종계의 품종에 따라 발생 병아리의 상태는 현저한 차이를 보이는데, 이는 품종 간 체형의 차이가 크고 종란의 무게 및 발생 병아리의 체중도 다를 뿐만 아니라, 유전적 특성에 따라 생존율의 차이도 큰 것으로 보고되고 있다(National Institute of Animal Science, 2011). 일반적으로 닭의 산란 연령이 증가함에 따라 난중이 증가하는데, 종계에 있어서도 연령이 증가할수록 종란이 무거워지고, 이에 따라 발생 병아리의 체중은 증가한다고 알려져 있다(Proudfoot et al., 1982; Wilson, 1991; Sklan et al., 2003; Ulmer-Franco et al., 2010; Alsobayel et al., 2013). 또한 산란 초기 때 생산한 병아리들의 폐사율이 상대적으로 높다고 보고하고 있는데(Wilson 1991, Peebles et al., 2004; Pedroso et al., 2005, Yassin et al., 2009), 특히 산란 초기의 종란은 난황낭(yolk sac)의 크기가 작아 영양성분이 부족하고 난황 지질과 난황 단백의 이용 능력이 떨어지기 때문이라고 하였다(Latour et al., 1998; Veira and Moran, 1998). 종란의 보관 기간이 발생 병아리에 미치는 영향에 대해서는 보관 기간이 길어지면 난질이 저하되어 병아리의 초기 생존능력에 부정적인 영향을 준다고 하였고(Tona et al., 2004; Fasenko, 2007; Yassin et al., 2009), 또한 부화 시간이 길어지게 되어 약추의 발생률을 증가시킨다고 하였다(Kirk et al., 1980; Christensen et al., 2002; Reijrink et al., 2010; Hamidu et al., 2011; Dymond et al., 2013).
발생 직후 병아리들의 강건성을 예측하기 위해서는 생존율 이외 초기 단계에서 병아리의 체중이나 사료 이용성과 같은 생산능력을 평가하거나 심장, 간, 비장과 같은 순환 및 면역 관련 장기 무게를 측정하여 이를 개체의 생리활성지표로 이용하고 있다(Puvadolpirod and Thaxton, 2000; Sekeroglu et al., 2011; Maatjens et al., 2014). 이 밖의 분자생물학적 표지로 interleukin(IL)과 같은 면역 관련 유전자의 활성도를 측정하거나(Niu et al., 2009; Helwig and Leon, 2011; Jang et al., 2014), telomere 길이를 측정하여 개체의 생리 활성 정도를 평가하고 있다(Sohn et al., 2012). 한편, 병아리의 스트레스에 대한 반응 정도는 열충격단백질(heat shock protein; HSP) 유전자 발현율이나 heterophil과 lymphocyte의 비율(H/L ratio), 혈장 코티코스테론 농도 및 DNA 손상률 등을 분석함으로써 개체의 스트레스 감수성 여부를 판단하고 있다(Lindquist and Craig, 1988; Sohn et al., 2015; Scanes 2016; Cho et al., 2020).
따라서 본 연구는 종계의 품종, 산란 연령 및 종란의 보관 기간이 병아리의 부화 후 강건성에 미치는 영향을 살펴보고자 한 것으로 한국 토종닭 종계 세 품종을 이용하여 산란 연령과 종란의 보관 기간에 따른 부화 시간, 발생 후 병아리의 생산능력 및 장기 무게를 조사하고, 이들 요인들이 개체의 생리적 활성도 및 스트레스 반응 정도에 미치는 영향을 분석하였다.
재료 및 방법
본 연구에서는 한국 토종닭인 황갈색 재래종(Hwanggalsaek Jaeraejong), 토착 로드아일랜드종(Korean Rhode Island Red) 및 토착 레그혼종(Korean White Leghorn) 세 품종을 시험 대상으로 하였다. 시험계들은 모두 암수 혼합(♀ 7:♂ 1) 형태로 평사에서 사육하였고, 이들이 생산한 종란은 경남과학기술대학교 종합농장 내 부화실에서 부화 후 발생시켰다. 종란은 종계의 산란 연령이 27~29주령 때와 50~52주령 때 수집하고, 각각 3일, 7일 및 14일간 보관 후 부화하였다. 수집한 종란은 각 시기별 1,080개씩 입란하였고, 두 기간에 발생한 병아리 중 1,400여 수를 분석 대상으로 하여 1주일간 사육하면서 분석 항목에 대한 조사를 수행하였다. 병아리들의 장기 표본 및 혈액 표본의 채취는 1주령 시험 종료 때 처리당 20수씩 무작위로 추출하여 분석에 공시하였다. 이러한 시험에 관련된 닭의 관리 및 취급은 본 대학 동물실험윤리위원회의 승인을 득한 후(IACUC Approved No. 2019-14) 제반 규정을 준수하여 시행하였다.
본 시험에 이용한 부화기는 발생좌와 발육좌가 구분된 5,000개 입란용 상업용 부화기(모델 BS-3240, 반석부화기, 시흥, 대한민국)로 발생좌의 온도는 37.8~38.3°C, 상대습도는 60%로 설정하였고, 발육좌의 온도는 37.8°C, 상대습도는 70%로 설정하였다. 입란은 품종과 보관 기간에 따라 처리 구 당 30개씩 4반복으로 배치하였다. 발생한 병아리들은 자동온도 및 환기 조절시스템이 완비된 육추사 내 케이지로 이송하고, 발생 후 1주일 간 사육하였다. 사육 온도는 최초 34°C에서 단계적으로 낮추어 시험 종료 때 32°C로 설정하였고, 사료 급여는 시중의 어린병아리 배합사료로 무제한 급여하였으며, 점등 관리는 23L:1D로 하였다.
부화 시간은 입란 시간부터 병아리가 부화된 시간으로 나타내었고, 병아리의 부화 기준은 난각을 완전히 깨고 나온 것으로 하여 2시간 간격으로 점검하였다. 부화 개시 때 종란의 무게를 측정하고, 부화 직후 병아리의 발생 체중과 7일령 체중을 측정하였다. 사료 섭취량은 7일간 처리 당 반복 구별로 사료 소비량을 측정하였다. 생존율은 병아리 입식 후 종료 때까지의 폐사를 매일 기록하여 입식 대비 생존 수로 계산하였다.
병아리들의 장기 표본은 시험 종료 때 처리별 20수씩 무작위로 선정하여 개체별 체중 측정 후 도살하였고, 간, 심장, 췌장 및 비장을 채취하여 각각의 무게를 측정하였다. 체중 대비 장기 비율은 도살한 개체의 생체중에 대한 장기의 무게 비율로 계산하였다.
Telomere 길이 및 HSP와 IL-6 유전자의 발현량은 Quantitative-PCR 방법을 이용하여 분석하였다(Cawthon, 2002; Cho et al., 2020). 시료의 준비는 각 개체의 혈액으로부터 RNA를 추출하고, 이들로부터 cDNA를 합성하였다. Real-time PCR을 위한 telomeric hexamer repeat(tel-1 & tel-2), HSP-70, HSP-90α, HSP-90β 및 IL-6 gene과 reference gene의 primers는 Table 1과 같다. Quantitative-PCR을 위한 반응물은 cDNA 9 μL(5 pmol/μL), SYBR Green(Roche, GmbH, Mannheim, Germany) 10 μL, primer(5 pmol/μL) 각각 0.5 μL를 넣어 최종 volume이 20 μL가 되도록 혼합하고, real-time PCR machine(Model LC480, Roche, Mannheim, Germany)을 이용하여 95°C에서 5분간 변성시킨 후 95°C 10초 변성, 60°C 30초 접합, 72°C 10초간 신장 반응을 60회 반복하면서 실시간 형광 모니터링하였다. Ct값의 측정은 LightCycler® 480 software v1.5(Roche Diagnostics, GmbH, Mannheim, Germany)를 이용하여 분석하였다. Reference gene을 이용한 각 분석 대상 유전자의 상대적 정량값은 Livak and Schmittgen(2001)이 제시한 2−ΔΔCt방법으로 측정하였다.
Heterophil과 lymphocyte의 비율(H/L ratio) 분석을 위하여 각 개체의 혈액 내 비율을 조사하였다(Jeong et al., 2020). 표본 제작은 채혈한 혈액을 슬라이드 글라스에 한 방울 떨어뜨리고 다른 슬라이드의 모서리를 이용하여 고르게 도말 후 건조하였다. 건조된 표본은 메탄올로 고정한 후 25% Giemsa stain solution(Sigma Chem, St Louis, MO, USA)으로 5분간 염색 처리하였다. 완성된 슬라이드는 광학현미경(Model BX-51, Olympus, Tokyo, Japan)으로 세포를 관찰하고 단위 면적당 heterophil과 lymphocyte 수를 측정하였다.
처리별 분석값에 대한 통계 모형은 고정모수모형(fixed model)으로 다음과 같다.
여기서, Yijkl = 각 관측 값
αi = i번째 품종의 효과
βj = j번째 생산연령의 효과
γk = k번째 보관 기간의 효과
(αβ)ij = i번째 품종과 j번째 생산연령 간의 상호작용 효과
(αγ)ik = i번째 품종과 k번째 보관 기간 간의 상호작용 효과
(βγ)jk = j번째 생산연령과 k번째 보관 기간 간의 상호작용 효과
(αβγ)ijk = i번째 품종과 j번째 생산연령 및 k번째 보관 기간 간의 상호작용 효과
εijkl = 실험 오차
종계의 품종, 산란 연령 및 종란의 보관 기간에 따른 병아리들의 처리 간 차이와 이들 처리 간 상호작용 효과는 SAS 통계 package(SAS Institite Inc., Cary, NC, USA)의 GLM procedure를 이용하여 분석하였다. 각 분석값에 대해 처리별 유의한 차이가 인정되었을 때는 동일 package의 Tukey’s HSD procedure로 평균값 간의 유의성 유무를 검정하였다.
결과 및 고찰
종계의 품종, 산란 연령 및 종란의 보관 기간이 발생 병아리의 생산 특성에 미치는 영향을 살펴보고자 입란 시 난중, 부화 시간, 발생 체중, 7일령 체중, 증체량, 사료 섭취량 및 생존율을 조사하였고, 이들의 결과를 Table 2와 3에 제시하였다.
부화 시간은 품종 및 보관 기간에 따라 유의한 차이를 보였는데(P<0.001), 황갈색 재래종이 토착 로드아일랜드종과 토착 레그혼종보다 약 7시간 정도 빨리 부화하는 것으로 나타났다. 또한 종란의 보관 기간이 길어질수록 부화 시간은 연장되는 양상을 보였다. 일반적으로 종란의 무게가 무거울수록 부화 시간이 길어진다고 하였는데(Ulmer-Franco et al., 2010), 본 연구에서도 입란 시 난중이 무거운 품종일수록 부화 시간이 길어지는 양상을 보였다. 산란 초기와 후기 간에 난중의 차이가 있어 난중이 가벼운 초기 종란의 부화 시간이 짧다는 보고가 있지만(Wilson et al., 1991; Sklan et al., 2003; Hulet et al., 2007), 종계의 산란 연령이 병아리의 부화 시간에 영향을 미치지 않는다는 보고도 있다(Hamidu et al., 2018). 본 연구에서는 산란 초기의 난중이 후기의 난중에 비해 유의하게 낮음에도 불구하고, 부화 시간의 차이는 없는 것으로 나타났다(Table 2, 3).
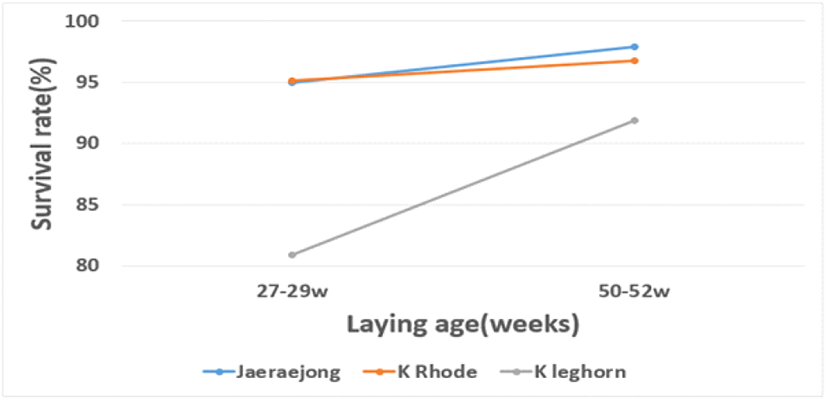
초생추의 생존율은 종계의 품종 및 산란 연령에 따라 영향을 받는 것으로 나타났는데, 토착 레그혼종이 다른 품종에 비해 낮은 생존율을 나타내었고, 산란 초기에 생산된 병아리들이 후기에 생산된 병아리에 비해 낮은 생존율을 나타내었다. 더불어 산란 연령에 따른 품종 간 생존율의 차이가 크게 나타나는데, 특히 토착 레그혼종의 산란 초기에 생산된 병아리들이 후기에 생산된 병아리에 비해 극히 낮은 생존율을 보였다(Fig. 1). 종계의 산란 연령에 따른 발생 병아리의 생존율의 차이는 많은 연구에서 어린 종계에서 생산된 병아리가 산란 후기 종계에서 생산된 병아리에 비하여 초기 폐사율이 높다고 보고하였다(Wilson, 1991; Peebles et al., 2004; Pedroso et al., 2005; Yassin et al., 2009). 이는 산란 초기 때 생산된 병아리가 산란 후기에 생산된 병아리에 비하여 체구가 작고 난황낭의 크기도 작아서 제공받는 영양 수준이 부족함으로 초기 폐사율에 영향을 미친다고 하였고(Viera and Moran, 1998), 어린 종계에서 생산된 병아리는 발육하는 동안 난황 지질 이용 능력이 떨어지고 지방 단백 이용 능력이 떨어지기 때문에, 초기 병아리의 폐사율이 증가한다고 하였다(Latour et al., 1998). 한편, 발생 초기 병아리의 성장과 품질은 난질의 영향을 받는데, 종란의 보관 기간이 길어지면 내부 난질이 저하되어 발생 병아리의 폐사율이 증가한다고 하였다(Tona et al., 2004; Fasenko, 2007; Yassin et al., 2009). 본 연구에서도 3일 보관된 종란에서 발생한 병아리의 생존율이 7일 이상 보관한 종란에서 발생한 병아리들에 비해 우수한 생존율을 보였다.
입란 시 난중은 품종, 종계의 연령 및 종란의 보관 기간에 따라 유의한 차이가 있었는데(P<0.001), 토착 레그혼종의 종란이 가장 무거웠고, 산란 후기의 종란이 초기 종란에 비해 무거웠으며, 보관 기간이 길어질수록 종란의 무게는 감소하였다. 병아리의 발생 체중은 입란 시 난중과 동일한 결과를 보였는데, 모든 처리구에서 공히 입란 시 종란 무게의 74~75% 정도로 나타났다. 그러나 7일령 체중의 경우, 발생 체중이 낮았던 처리구의 병아리들의 증체 효과가 두드러지게 나타났는데, 황갈색 재래종의 증체량 및 사료 섭취량이 다른 두 품종에 비해 유의하게 높게 나타났고(P<0.01), 산란 초기에 생산된 병아리들이 후기 생산 병아리에 비해 높은 증체량과 더불어 7일령 체중에서 오히려 발생 체중과 역전되는 결과를 보였다. 그러나 종란의 보관 기간에 따른 증체량, 사료 섭취량 및 7일령 체중의 차이는 없는 것으로 나타났다. 이와 같은 결과는 어린 종계에서 생산된 병아리에 비해 나이가 많은 종계에서 생산된 병아리의 발생 체중은 높지만, 후기 증체량은 어린 종계의 병아리가 훨씬 양호하였다는 보고나, 브로일러에서 산란 초기에 생산된 병아리의 체중이 후기 생산 병아리보다 더 가벼웠으나 출하 체중에는 차이가 없었다는 보고와 거의 일치하는 양상이다(Hulet et al., 2007; Ulmer-Franco et al., 2010). 이러한 현상은 개체의 보상 성장 효과에 기인한 결과로서 발생 시 낮은 체중에 따른 부족한 영양소 보충을 위하여 사료 섭취량의 증가와 더불어 증체량이 증가하였기 때문이다.
병아리의 생리 활성 상태를 파악하기 위하여 종계의 품종, 산란 연령 및 종란의 보관 기간에 따른 발생 병아리들의 장기 무게를 측정하였다. 분석 결과, 처리별 7일령 병아리들의 간, 심장, 췌장 및 비장의 무게를 Table 4에 제시하였고, 이들의 체중 대비 장기의 무게 비율을 Table 5에 제시하였다.
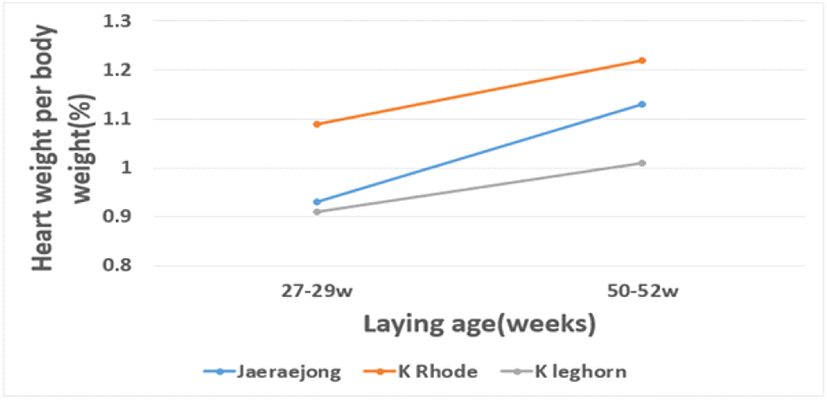
췌장의 생체중 대비 장기 비율을 제외한 모든 조사된 장기의 무게나 장기의 비율은 품종 간에 유의한 차이를 보이는데(P<0.01), 공시 품종 중 토착 레그혼종의 장기 무게가 다른 종에 비해 다소 무거운 양상을 나타내었다. 그러나 이들 장기 중 심장의 무게나 생체중 대비 심장 무게의 비율은 토착 레그혼종이 다른 종에 비해 유의하게 낮은 결과를 보이며, 특히 산란 후기에 생산된 병아리에서 더욱 낮게 나타났다(Fig. 2). 산란 연령에 따른 발생 병아리의 장기 무게나 장기의 비율은 간을 제외하고 모든 장기의 무게나 비율이 산란 초기에 생산된 병아리가 산란 후기에 생산된 병아리보다 낮은 것으로 나타났다. 종란의 보관 기간에 따라서는 3일 보관 후 발생한 병아리가 7일 이상 보관 후 발생한 병아리에 비해 간, 심장, 췌장의 무게가 더 무거운 결과를 보였으나, 생체중 대비 장기 무게의 비율에서는 간을 제외한 모든 장기에서 보관 기간 간 차이가 없는 것으로 나타났다. 대부분의 장기 무게는 생체중과 비례한 중량비를 나타내고 있는데, 심장의 무게와 비율은 생체중 무게와 관련이 없는 것으로 나타났다. 심장은 체내의 혈액순환을 담당하는 중요한 장기인데, 심장의 크기가 작다는 것은 혈액이 필요로 하는 장기에 혈액 공급이 원활히 이루어지지 못함을 의미한다(Maatjens et al., 2014). 간은 소화 효소의 분비 외 면역 염증 반응에 대한 대사 작용을 담당하는 기관으로 이의 크기는 이들 대사 활성 정도를 간접적으로 나타내는 지표이다(Pearson et al., 1983; Puvadolpirod and Thaxton, 2000). 비장 또한 면역 관련 기관으로 체내의 스트레스 반응이 많아지면 이의 크기가 상대적으로 줄어드는 것으로 알려져 있다(Pope, 1991; Ravindan et al., 2006). 토착 레그혼종의 경우, 심장의 크기와 비율이 체구에 비해 상대적으로 작은 것으로 나타났는데, 이는 강건성의 저하를 시사하는 것으로 작은 심장은 체내 혈액 공급을 원활하게 하지 못하므로 심장의 운동량을 증가시켜 과부하를 야기하고, 이로 인해 내부 장기의 역할을 저해하게 된다. 따라서 토착 레그혼종의 초기 생존율 저하는 이들의 작은 심장과 다소 관련이 있는 것으로 보인다. 더불어 산란 초기 때 생산한 병아리가 후기에 생산한 병아리에 비해 낮은 생존율을 보였는데, 이 또한 산란 초기 병아리의 작은 심장 크기와 관련이 있는 것으로 생각된다. 병아리의 강건성과 관련하여 종란의 보관 기간이 병아리의 장기 무게에 미치는 영향은 거의 없는 것으로 판단된다.
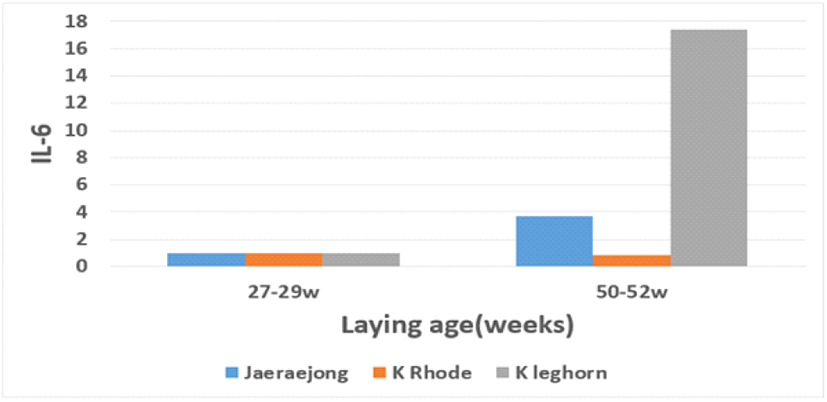
종계의 품종, 산란 시 연령 및 종란의 보관 기간이 발생 병아리의 면역 및 생리 활성도에 미치는 영향을 살펴보고자 IL-6의 유전자 발현 수준 및 텔로미어의 상대적 길이를 분석하였고, 이의 결과를 Table 6에 제시하였다. 분석 결과, 병아리의 IL-6 유전자 발현율에 있어 품종 및 산란 연령 간의 차이를 보이고, 종란의 보관 기간에 따른 차이는 없었다. 품종 간에는 토착 로드아일랜드종의 병아리가 황갈색 재래종과 토착 레그혼종에 비해 낮은 발현 수준을 나타내었고, 산란 연령에 따라서는 산란 후기에 생산된 병아리가 초기에 생산된 병아리에 비해 높은 발현 수준을 나타내었다. 특히 산란 후기에 생산한 토착 레그혼종 병아리의 IL-6 발현 수준이 유의하게 높은 양상을 보였다(Fig. 3). IL-6는 대식세포에서 분비되는 친염증성 cytokine 중 하나로서 조혈계와 신경계 세포의 증식과 분화에 관여하고, 면역 반응 때 간세포에 작용하여 급성기 단백질의 합성을 유도한다. 일반적으로 체내 면역 이상이나 염증성 질환 발생 시 IL-6의 과발현이 나타나는 것으로 알려져 있다(Van Snick, 1990). 본 연구에서 산란 후기에 생산한 토착 레그혼종 병아리의 IL-6 발현 수준이 높다는 것은 이들의 체내 염증반응이 다른 종에 비해 증가함을 시사한다. 한편, 텔로미어의 상대적 길이 분석 결과, 품종 및 종란의 보관 일수에 따른 차이는 없었고, 산란 초기에 생산된 병아리가 후기에 생산된 병아리에 비해 유의하게 긴 텔로미어 길이를 가진 것으로 나타났다. 텔로미어는 염색체 말단부를 지칭하는 것으로 세포 분열이 진행됨에 따라 텔로미어의 길이는 짧아지게 되므로 이를 대표적인 노화의 표지로 간주한다(Harley et al., 1990, 1992). 그러나 텔로미어의 감축 정도나 속도는 다양한 요인들에 의해 영향을 받는데, 품종 및 계통과 같은 유전적 요인뿐만 아니라, 환경적 요인에 의해서도 큰 영향을 받는 것으로 알려져 있다(Richter and Proctor, 2007; Sohn and Subramani, 2014). 따라서 텔로미어의 길이나 감축 정도를 개체의 생명 활성도 및 스트레스 반응 정도의 간접 표지로 이용한다(Sohn et al., 2012). 본 연구 결과, 산란 초기에 생산된 병아리의 텔로미어 길이가 후기에 생산된 병아리보다 길다는 것은 모체의 텔로미어 길이가 자손에게 영향을 미친 것으로 생각할 수 있으나, 이에 대해서는 많은 논란이 있다. 초기 체세포 복제동물에서 공여 세포의 연령이 수정란의 텔로미어 길이에 영향을 준다는 보고(Shiels et al., 1999)가 있는 반면, 공여 세포 연령과 관계없이 수정란의 배반포 단계에서 텔로미어 길이는 reprogramming이 되어 발생 시 일정 길이로 신장한다는 보고도 있다(Lanza et al., 2000). 닭의 경우 모계 연령에 따른 생산 배아의 텔로미어 함량 비교 분석에서 모계의 산란 연령과 배아의 텔로미어 함량 간에는 거의 상관관계가 없는 것으로 보고하였다(Lee et al., 2009; Park and Sohn, 2014). 비록 산란 초기에 생산된 병아리들이 후기에 생산된 병아리에 비해 생존율이 낮고 발생 시 체중은 낮았지만, 7일령 체중에서는 역전 양상을 보이고 더불어 긴 텔로미어 길이와 낮은 IL-6 발현 수준은 이들의 생리활성 정도가 산란 후기 병아리에 비해 상대적으로 높다는 것을 시사한다.
종계의 품종, 산란 시 연령 및 종란의 보관 기간에 따른 병아리의 체내 스트레스 반응 정도를 살펴보고자 처리 개체별 HSP-70, HSP-90α, HSP-90β의 유전자 발현 수준 및 H/L ratio를 분석하였고, 이의 결과를 Table 7에 제시하였다. 분석 결과, HSP 유전자의 발현 수준 및 H/L ratio 값이 토착 레그혼종 병아리들이 다른 품종에 비해 상대적으로 높게 나타났고, 산란 후기에 생산된 병아리가 산란 초기 생산 병아리에 비해 높게 나타났다. 반면, 종란의 보관 기간이 병아리들의 HSP 발현 수준 및 H/L ratio 값에 미치는 영향은 거의 없는 것으로 분석되었다. Heat Shock Proteins는 열 스트레스에 반응하여 합성되는 특이 단백질 군으로서 열 스트레스를 받은 세포의 경우 HSP들이 열 민감 단백질들과 결합하여 세포를 보호하는 역할을 하므로 이의 발현율이 증가하게 된다(Lindquist, 1986; Schlesinger, 1990). 닭의 경우 열 스트레스뿐만 아니라, 외부 스트레스 적 환경에 노출되었을 때도 HSP의 발현율은 상승한다고 보고하였다(Zulkifli et al., 2002; Beloor et al., 2010; Sohn et al., 2012). H/L ratio는 heterophil과 lymphocyte의 비율로 스트레스로 인한 염증반응이 발생하면 heterophil이 증가하게 되고, lymphocyte의 비율은 감소하게 되어 H/L ratio가 증가하게 된다(Gross and Siegel 1983; Gross 1989; McFarlane and Curtis 1989). 따라서 이들의 분석 결과 토착 레그혼종의 병아리가 다른 품종들에 비해서 스트레스에 더욱 민감하게 반응하는 것으로 보이고, 산란 후기에 생산된 병아리가 산란 초기에 생산된 병아리에 비해 스트레스 반응이 더욱 민감하다고 생각된다. 한편, 토착 로드아일랜드종 병아리들의 HSP발현 수준이 다른 품종에 비해 상대적으로 낮게 나타나 스트레스 민감도가 낮은 것으로 판단되는데, 이는 한국 토종닭 품종들의 스트레스 반응 정도 비교에 관한 연구에서 토착 로드아일랜드종의 스트레스 민감도가 가장 낮았다는 보고와 잘 일치되는 결과이다(Sohn et al., 2015; Cho et al., 2020).
이상의 결과들을 종합하면, 종계의 품종과 산란 시 연령이 발생 병아리의 강건성에 영향을 미치는 것으로 보이고, 종란의 보관 기간은 병아리의 강건성에 크게 영향을 미치지 않는 것으로 생각된다. 공시된 세 품종의 토종닭 중 토착 레그혼종 병아리들의 강건성이 가장 약한 것으로 보이는데, 이는 생존율이 저조하고 상대적으로 작은 심장을 가지며 모든 생리활성지표가 낮고 스트레스 반응 지표들이 높았기 때문이다. 또한 산란 초기에 생산한 병아리들이 산란 후기에 생산한 병아리에 비해 생존율은 낮았지만, 생존 개체들의 생리활성지표가 높고, 스트레스 반응 지표들이 낮아 상대적으로 강건성이 높다고 판단된다.