서 론
발생 직후 병아리의 암수 감별은 양계산업의 경제적 관점에서 매우 중요한 요소이다. 그러므로 오래전부터 병아리의 암수를 감별하고자 하는 다양한 시도가 있었고, 이러한 시도들 중 지금까지 가장 대표적인 성 판별 방법은 발생 병아리의 내부 생식기의 형태적 차이로써 성을 판별하는 항문돌기감별법으로 알려져 있다(Masui et al., 1925; Martin, 1934). 그러나 이러한 판별법은 감별기술을 숙련하는 데 오랜 시간이 필요하고, 또한 전문 감별사에 의존하여야 하므로 시간적, 경제적 부담이 큰 단점이 있다. 따라서 최근에는 닭의 반성유전(sex-linked inheritance)의 원리를 이용한 병아리의 자가성감별법(auto-sexing)이 산업적으로 널리 이용되고 있다. 최근에는 부화 전 배자 상태에서 병아리의 성을 판별하고자 하는 연구들이 활발히 시도되고 있지만, 아직은 이러한 병아리의 성 판별 기법들이 기술적, 경제적인 문제들로 인해 상업적으로 적용하기에는 많은 어려움이 있다(Steiner et al., 2011; Weissmann et al., 2013; Webster et al., 2015; Clinton et al., 2016; Galli et al., 2018; Alin et al., 2019). 반성유전이란 성 염색체상에 성 이외의 형질을 발현하는 유전자가 성염색체와 연관되어 유전되는 현상을 일컫는 것으로 이러한 유전자를 반성유전자(sex-linked gene)라 하고, 닭에 있어 깃털의 줄무늬 띠를 형성하는 횡반유전자(Spillman, 1908), 깃털 성장 발달을 지연하는 만우성 유전자(Serebrovsky, 1922), 갈색 등 유색에 대해 우성인 은색유전자(Henderson, 1959) 등이 깃털과 관련된 대표적인 반성유전자들이다. 특히, 이들 중 닭의 깃털 발육 속도와 연관된 만우성 유전자를 이용한 병아리의 성 감별법이 상업용 실용계에 가장 보편적으로 이용되고 있다(Bitgood, 1999; Wilson et al., 2007; Aviagen Brand 2017). 만우성(late-feathering)이란 깃털의 발육 속도가 느린 성질을 의미하는 것으로, 깃털의 발육 속도가 빠른 조우성(early-feathering)에 대해 우성이며 멘델 유전 양식에 따라 유전된다. 따라서 깃털 조만성유전자를 이용한 병아리의 성 감별법의 원리는 어미 닭을 만우성(ZKW)으로 하고, 아비 닭을 조우성(ZkZk)으로 하였을 때 수평아리는 만우성, 암평아리는 조우성이 되어 깃털 발육의 형태로써 병아리의 성을 판별하는 것이다(Somes, 1969; Iraqi and Smith, 1995; Elferink et al., 2008). 그러므로 만우성 유전자를 이용한 병아리의 성 감별이 산업적으로 실용화되기 위해서는 종계의 기초 계군 내 조우성과 만우성 유전자가 존재하여야 하고, 깃털 조만성에 따른 개체 간 생산능력의 차이도 없어야 하며, 종계 계통 조성 후 생산되는 병아리의 성 판별이 정확하여야 한다. 국내 상업용 실용계 종계의 대부분은 외국의 글로벌 종계회사로부터 수입하고 있다. 이들 실용계의 상당수는 깃털 감별법으로 암수를 감별하고 있는데, 이는 수입 종계들이 이미 자가성감별 계통으로 부계와 모계가 조성된 상태이기 때문이다. 그러나 한국토종닭의 경우, 아직 깃털 감별법을 이용한 병아리의 암수 감별이 산업적으로 활용되지 않고 있다. 이는 산업적 수요가 제한적이고 이에 따른 연구도 활발하게 이루어지지 않았기 때문이다. 그러나 다행스러운 것은 이러한 연구에서 한국토종닭 집단에도 조우성과 만우성 유전자가 함께 분포하고, 조우성과 만우성 개체 간에 생산능력의 차이가 없다고 보고함으로써 깃털 감별의 가능성을 시사한 것이다(Sohn et al., 2012, 2013; Bang et al., 2018; Kim et al., 2019).
따라서 본 연구는 한국토종닭의 깃털 감별의 산업적 적용을 위하여 닭의 조우성과 만우성 유전자를 이용한 자가성감별 계통을 조성하고자 한 것으로 한국토종닭 대상 집단 내 조우성과 만우성 개체의 분포 현황 및 유전자 빈도를 추정하고, 이를 바탕으로 조우성 부 계통과 만우성 모 계통의 종계 집단을 조성한 후, 조우성 종계와 만우성 종계 간의 생산능력을 비교 검토함과 동시에 조성된 자가성감별 계통으로부터 생산된 병아리들의 깃털 감별의 유효성을 검정하였다.
재료 및 방법
본 연구에 이용된 한국토종닭 기초 계군은 경남과학기술대학교 종합농장에서 사육 중인 한국재래닭 적갈색종 557수를 대상으로 하였다. 시험계들은 본 대학 종합농장 내 부화사에서 자체 부화하여 발생한 개체로서 14주령까지 강제 환기와 자동 급이 및 자동온도조절시스템이 완비된 배터리형 케이지(660 cm2/1수)에서 사육하고, 이후 40주령까지는 군사형 케이지(1,157 cm2/수) 및 평사식 종계사(2,370 cm2/1수)에서 암수 혼합 사육하였다. 사료 급여는 사육단계별로 어린병아리, 중병아리, 산란전기, 산란초기 및 산란중기로 나누어 상업용 시판 사료로 자유 급여하였다. 점등관리는 점감점증법으로 발생 후 16주령까지 점감하고, 이후 26주령까지 점증 후 사육 종료 때까지 16시간으로 고정하였다. 백신접종 등 그 밖의 일반 닭 사양관리는 경남과학기술대학교 닭 사육관리 지침에 따라 이루어졌고, 시험에 관련된 닭의 관리 및 취급은 본 대학 동물실험윤리위원회(IACUC)의 승인을 득한 후 규정을 준수하여 시행하였다.
깃털 형태에 의한 조만성 식별은 부화 직후 날개깃의 형태로써 감별하였다. 식별 방법은 Bang et al.(2018)이 제시한 병아리의 깃털 발생 양상을 기준으로 날개깃의 주익우(primaries)와 부익우(coverts)의 형태와 길이의 차이로 구분하였다. 주익우가 평균 13 mm 정도로 길고, 부익우의 길이가 주익우의 절반 정도인 개체를 조우성으로 분류하고, 주익우가 8 mm 정도로 짧고 주익우와 부익우의 길이 차이가 거의 없는 개체는 만우성으로 분류하였다.
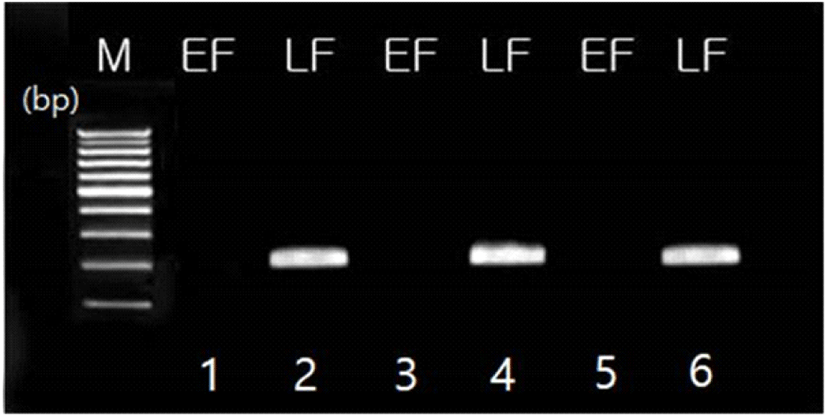
조우성 개체와 만우성 개체의 식별을 위한 분자생물학적 검증으로 만우성 특이 DNA primer를 이용하여 PCR을 수행하였다. 시료로 사용할 genomic DNA는 혈액으로부터 추출하였고, 만우성 특이 DNA primer는 Iraqi and Smith(1994)가 제시한 JFIL-1 유전자의 일부 영역을 대상으로 제작하였다. PCR은 TaKaRa TaqTM kit(Takara, Kyoto, Japan)를 이용하여 10 × buffer 2.5 μL, dNTP 2 μL, Taq polymerase(0.5 unit/μL) 2 μL, primer 4 μL(forward 2 μL, reverse 2 μL)와 genomic DNA 3 μL(100 ng/μL), ddH2O 11.5 μL를 넣어 총량이 25 μL가 되도록 혼합한 후 PCR machine(Takara, Kyoto, Japan)으로 증폭시켰다. PCR 반응 조건은 95°C에서 5분간 최초 변성시키고, 95°C에서 1분간 변성, 56°C에서 1분간 접합, 72°C에서 2분간 신장하는 3단계의 과정을 30회 반복한 후 72°C에서 15분간 마지막 신장 과정을 실시하였다. 반응물은 1.5% agarose gel에서 125 V로 15분간 전기영동한 후 만우성 유전자의 band 유무를 관찰하였다.
조우성과 만우성 개체 간 생산능력의 비교는 생존율, 체중, 초산일령, 산란율 및 난중에 대해서 발생 이후 40주령까지 각 형질을 측정하고 비교 분석하였다. 생존율은 조우성과 만우성의 식별이 끝난 2주령부터 40주령까지 시험개시일의 개체 수에 대한 생존 수수의 비율로 나타내었다. 체중은 부화 직후부터 10주령 단위로 개체별 체중을 측정하였다. 초산일령은 각 계군의 산란율이 5%에 도달하는 날을 기준으로 산출하였고, 산란율은 21주령부터 40주령까지 주령별 생존 수수 대비 총 산란수의 비율로 일계산란율(hen-day egg production)을 산출하였다. 난중은 계군별로 150일령 및 270일령에 생산한 달걀 전수를 대상으로 무게를 측정하였다.
만우성 수컷 개체가 동형접합체(KK)인지 이형접합체(Kk)인지를 알기 위하여 열성인 조우성 암컷 개체(k-)와 검정교배를 수행하였다. 검정 대상은 만우성 수컷 40수로 이들의 유전자형을 확인하기 위하여 수컷 1수당 조우성 암컷 6수를 교배시키고, 이들로부터 생산된 자손들의 조우성과 만우성의 분포 비율을 조사하였다.
결과 및 고찰
한국재래닭 적갈색종 557수를 대상으로 기초 집단 내 깃털 형태에 따른 조우성과 만우성 개체를 식별하였고(Fig. 1), 조우성과 만우성 개체의 분포 현황과 유전자 및 유전자형 빈도를 추정하여 이의 결과를 Table 1에 제시하였다. 조사 결과, 본 집단 내 조우성 개체가 429수(77%), 만우성 개체가 128수(23%)로 확인되어 만우성보다 조우성 개체의 빈도가 높게 나타났다. 또한 본 집단이 Hardy-Weinberg 평형상태에 있다는 가정하에 유전자 빈도를 추정한 결과, 수컷의 조우성유전자(k) 빈도는 0.784, 만우성 유전자(K) 빈도는 0.216으로 추정된 반면, 암컷에서는 조우성유전자 빈도가 0.824, 만우성유전자 빈도가 0.176으로 추정되어 수컷 집단 내 만우성유전자 빈도가 암컷 집단에 비해 다소 높게 나타났다. 이를 토대로 본 집단 내 전체 조만성유전자 빈도를 계산한 결과, 조우성유전자(k)의 빈도는 0.814, 만우성유전자(K)의 빈도는 0.186으로 추정되었다. 국내에서 사육하는 품종들의 집단 내 깃털 조만성의 분포 양상과 유전자 빈도에 관한 연구에서 Sohn et al.(2012)은 국가 보존 유전자원 토종닭 10품종을 대상으로 깃털 조만성의 분포 양상을 조사한 결과, 토종 흑색코니시종, 토종 로드아일랜드레드종 및 한국재래닭 적갈색종에만 조우성과 만우성 개체들이 혼재되어 있고, 토종 갈색코니시종, 백색레그혼종, 오골종, 재래황갈종, 재래회색종, 재래백색종 및 재래흑색종은 집단 내 조우성 개체만 존재한다고 보고하였다. 더불어 한국재래닭 적갈색종의 집단 내 조우성유전자 빈도는 0.856∼0.871, 만우성유전자 빈도는 0.129∼0.144로 보고하였다. 또한 Bang et al.(2018)은 ㈜한협원종이 보유한 토종 종계로부터 생산된 실용 토종닭 병아리 856수를 대상으로 깃털 조만성 분석 결과, 조우성이 744수이고, 만우성이 112수로 나타나, 이들의 종계 집단 내 조우성과 만우성유전자가 함께 있음을 제시하였다. 이처럼 집단 내 조우성과 만우성 개체의 빈도는 품종에 따라 큰 차이를 보이고 있고, 조우성과 만우성 개체가 함께 존재하는 계군의 집단은 많지 않은 것으로 나타났다. 깃털 자가성감별 계통 조성을 위해서는 반드시 집단 내 조우성과 만우성유전자가 혼재되어 있어야 하고, 이들 유전자의 분포 빈도가 일정 수준은 유지되어야 한다. 따라서 본 연구의 한국재래닭 적갈색종 기초 집단의 경우 조우성과 만우성 개체가 함께 있고, 조우성과 만우성 유전자 빈도 또한 깃털 자가성감별 계통 조성을 위한 가능한 빈도로 나타났다.

조우성과 만우성유전자를 이용하여 병아리의 자가성감별을 하기 위해서 모계는 만우성(ZKW)으로, 부계는 조우성(ZkZk)으로 고정되어야 한다. 따라서 자가성감별을 위한 종계의 모계통 조성을 위한 원종계의 교배조합은 ZKZK⨯ ZKW로 하는 것이 바람직하다. 그러므로 모계통의 조성을 위해 만우성 유전자를 동형접합체(KK)로 가진 수컷의 선발이 필수적 요소이다. 우성 형질에 있어 특정 개체가 동형접합체를 가졌는지, 이형접합체를 가졌는지는 열성동형접합체를 가진 개체와 검정교배하여 생산된 자손들의 형질 분리비로 쉽게 판별할 수 있다. 본 실험에서는 우성 형질인 만우성으로 확인된 한국재래닭 적갈색종 수컷 40수의 유전자형 검정을 위해 열성 형질인 조우성 암컷 개체와 수당 6수씩 검정 교배하여 생산된 자손들에 대한 조우성과 만우성의 분리비를 살펴보았다. 분석 결과, 자손 중 1마리라도 조우성 개체를 생산한 검정 수컷은 37수이고, 모두 만우성 자손만을 생산한 검정 수컷은 3수였다. 따라서 검정 수컷 중 조우성 자손이 한 마리도 생산되지 않은 3마리의 수컷을 만우성 동형접합체(ZKZK)를 가진 개체로 추정하였고, 나머지 37수는 이형접합체 형태의 만우성 개체(ZKZk)로 판단하였다. 이러한 검정교배로부터 생산된 총 420수의 자손에 대한 깃털 조만성의 발생 분포 양상은 Table 2와 같다.
| Crossings | EF progeny | LF progeny | ||||
|---|---|---|---|---|---|---|
| ♂ | ♀ | Total | ♂ | ♀ | Total | |
| Number of chicks | ||||||
| ♂ Heterozygous LF(ZKZk) x ♀ EF(ZkW) | 62 | 56 | 118 | 119 | 135 | 254 |
| ♂ Homozygous LF(ZKZK) x ♀ EF(ZkW) | 0 | 0 | 0 | 24 | 24 | 48 |
조우성과 만우성 개체 간 생산능력의 차이는 자가성감별 계통을 조성하는 데 매우 중요한 요인으로 작용한다. 따라서 기초 집단 내 조우성 개체와 만우성 개체 간 생산능력을 비교하기 위하여 만우성 개체 128수와 조우성 개체 429수에 대한 생존율, 산육능력 및 산란능력을 비교 분석하였다. Table 3은 2주령부터 40주령까지 시험계들의 생존율을 나타낸 것으로 2∼14주령까지 육성기와 15∼40주령까지 산란기로 나누어 분석하였다. 분석 결과, 시험 전 기간 동안 조우성 집단이 만우성 집단에 비해 높은 생존율을 나타내었고, 특히 산란기 동안 두 집단 간 생존율의 유의한 차이를 보였다(P<0.01). 깃털의 조만성이 생존율에 미치는 영향에 대해서는 다양한 결과들을 제시하고 있는데, 만우성 유전자는 ev21 유전자와 연관되어 있으므로 leukosis virus의 감염에 취약하여 조우성 개체보다 생존율이 낮다고 하였다(Harris et al., 1984; Bacon et al., 1988; Smith and Fadly, 1994). 반면 깃털 조만성에 따른 마렉병의 감염율이나 폐사율의 차이는 없었으며, 브로일러에서도 조우성 개체와 만우성 개체 간 생존율의 차이는 없다고 하였다(Some and Jakowski, 1974; Lowe and Merkley, 1986). 한편, 한국재래닭을 대상으로 깃털 조만성에 따른 생존율의 비교 연구에서 발생 이후 60주령까지 시험 전 기간에 걸쳐 조우성과 만우성 개체 간 생존율의 차이는 없다고 하였고(Sohn et al., 2013), 토종 종계에서는 암수 모두 만우성 집단이 조우성 집단보다 생존율이 우수하다고 보고하였다(Kim et al., 2019). 이와 같이 한국토종닭 집단에서 깃털 조만성이 생존율에 미치는 영향에 대해서는 각기 다른 결과들을 제시하고 있다. 따라서 생존율은 시험계들의 사양관리적 환경이 주된 영향을 미치는 것으로 생각되고, 깃털 조만성에 의한 영향은 거의 없는 것으로 사료된다.
| Feathering types | Growing period (2~14wks) | Laying period (15~40wks) | Total period (2~40wks) |
|---|---|---|---|
| % | |||
| EF chickens | 100±0.0 | 97.7±1.3a | 98.6±2.3a |
| LF chickens | 99.2±0.4 | 94.1±3.2b | 94.5±4.9b |
깃털 조만성에 따른 산육능력의 차이를 살펴보고자 조우성 개체와 만우성 개체들의 발생시, 10주령, 20주령, 30주령 및 40주령 체중을 측정하고, 이들 간의 차이를 비교 분석하여 이의 결과를 Table 4에 제시하였다. 분석 결과, 시험 전 기간 동안 조우성 집단과 만우성 집단 간 수컷 체중의 차이는 없었고, 암컷은 발생 시 체중과 40주령 체중을 제외한 모든 주령에서 차이가 없었다. 깃털 조만성이 산육능력에 미치는 영향에 대한 보고에서 브로일러의 경우 조우성 개체들이 만우성 개체에 비해 빠른 성장과 높은 체중을 보인다고 보고(Dunnington and Siegel, 1986; Khosravinia, 2008)한 반면, 만우성 개체의 성장 능력이 더 우수하다는 보고(Lowe and Merkley, 1986; Merkley and Lowe, 1988; Nahashon et al., 2004)도 있으며, 산육능력과 깃털 조만성 간에 관련이 없다는 보고도 있다(O'Sullivan et al., 1991; Fotsa et al., 2001). 한국재래닭의 경우, 발생 후 60주령까지 모든 주령에서 조우성 개체와 만우성 개체 간 체중의 차이는 없는 것으로 보고하였고(Sohn et al., 2013), 토종 종계에서도 사육 기간 전반에 걸쳐 조우성과 만우성 암수 모두 체중의 차이는 없다고 보고하였다(Kim et al., 2019). 따라서 본 결과와 더불어 한국재래닭에 있어 깃털의 조만성과 산육능력 간에는 거의 관련성이 없는 것으로 판단된다.
조우성 개체와 만우성 개체 간 산란능력을 비교하고자 초산일령, 난중 및 산란율을 조사하고, 이의 결과를 Table 5에 제시하였다. 분석 결과, 초산일령은 조우성 집단이 만우성 집단보다 약 6일 정도 빠른 것으로 나타났으나, 일계산란율은 만우성 집단이 조우성 집단보다 유의하게 높은 것으로 나타났다(P<0.05). 반면, 150일령 및 270일령에 산란한 달걀의 무게는 두 집단 간 차이가 없는 것으로 나타났다. 산란계의 경우, 조우성 개체가 만우성 개체보다 초산일령이 빠르고 산란능력이 우수하다고 보고하였고(Somes, 1970; Havenstein et al., 1989; Durmus et al., 2010; Goger et al., 2017), 겸용종 또는 육용 종계의 경우는 깃털 조만성에 따른 산란능력의 차이는 없다고 하였다(Ledvinka et al., 2011, 2014; Mincheva et al., 2012). 한편, 한국재래닭의 경우, 조우성 개체들이 만우성 개체들에 비해 빠른 초산을 보이지만, 조우성과 만우성 개체 간의 산란능력의 차이는 없다고 하였고(Sohn et al., 2013), 토종 종계의 경우 조우성과 만우성 개체 간 초산일령의 차이는 없었으나, 만우성 개체들이 조우성 개체들보다 산란율이 높았다고 보고하였다(Kim et al., 2019). 난중은 산란계에 있어 만우성 개체의 달걀이 조우성 개체의 달걀에 비해 다소 무거웠다고 하였으나(Lowe et al., 1965; Somes, 1970), 여러 겸용종에서는 조만성 개체 간 난중의 차이는 없다고 하였다(Lowe and Garwood, 1981; Mincheva et al., 2012; Ledvinka et al., 2014; Goger et al., 2017). 한편, 한국재래닭 및 토종 종계에 있어서도 깃털 조만성에 따른 난중의 차이는 없다고 하였다(Sohn et al., 2013; Kim et al., 2019). 이와 같이 연구자에 따라 조우성과 만우성 개체 간 산란능력의 결과를 다소 달리 제시하고 있으나, 한국재래닭의 경우 깃털 조만성에 따른 산란능력의 차이는 없거나, 만우성 개체들이 조우성 개체보다 좀 더 우수한 것으로 나타났다. 한국토종닭에 있어 만우성 개체들의 산란능력이 조우성 개체에 비해 우수하다는 것은 자가성감별 모계통으로 만우성 개체를 이용하여야 하기 때문에 매우 바람직한 결과로 사료된다.
깃털 조만성의 유전적 전이 양상을 살펴보고자 한국재래닭 적갈색종을 이용하여 깃털 양상에 따라 4가지 조합의 교배조합 검정시험을 실시하였다. 교배조합의 구성은 만우성 ♂ × 만우성 ♀, 조우성 ♂ × 조우성 ♀, 만우성 ♂ × 조우성 ♀ 및 조우성 ♂ × 만우성 ♀으로 하여 각 교배조합별 생산된 병아리의 깃털 조만성 양상을 확인하고, 이들의 발생 수와 분포 빈도를 Table 6에 제시하였다. 시험 결과, 만우성 ♂ × 만우성 ♀의 경우, 생산된 병아리 암수 모두에서 조우성과 만우성이 나타났다. 이는 만우성 수컷(ZKZK, ZKZk)과 만우성 암컷(ZKW) 간의 교잡에서 이론적으로 조우성 수컷 병아리(ZkZk)는 생산될 수 없는데, 본 교잡 결과 3수의 조우성 수컷 병아리가 생산되어 이론치와 다른 양상을 보였다. 이러한 원인은 교잡 시험에 공시된 만우성 어미 중 최소 한 수 이상이 조우성이었기 때문으로 판단되는데, 이는 종계의 조만성 구분 오류로 생각된다. 그러나 조우성 ♂ × 조우성 ♀의 경우는 교배 시험 결과 생산된 병아리가 모두 조우성 개체로 나타나, 이론치와 100% 일치하는 결과를 보였다. 한편, 만우성 ♂ × 조우성 ♀의 경우, 생산된 암수 병아리 모두에서 조우성과 만우성이 나타났고, 조만성 비율은 조우성이 36.2%, 만우성이 63.8%로 나타났다. 이는 이론적으로 만우성 수컷(ZKZK, ZKZK)과 조우성 암컷(ZkW) 간의 교잡에서 조우성 병아리와 만우성 병아리가 암수 모두 생산될 수 있고, 병아리의 조만성의 분포 비율은 집단 내 만우성 유전자의 빈도에 따라 달라진다. 마지막으로 조우성 ♂ × 만우성 ♀의 경우는 깃털 자가성감별에 이용되는 교배조합으로 이론적으로 암평아리는 모두 조우성, 수평아리는 모두 만우성으로 생산된다. 본 실험 결과, 만우성의 암평아리는 생산되지 않았지만 2수의 조우성 수컷이 생산되어 이론적 분리비와 다른 양상을 보였는데, 이는 병아리의 조만성 깃털의 식별 오류로 PCR 검정 결과 확인되었다(Fig. 2). 상기 교배조합 시험에 따른 자손의 조만성 전이 양상이 이론적인 분리비와 일치하는가를 카이자승법(chi-square test)으로 검정하여 이의 적합도와 유효성을 Table 7에 제시하였다. Table 7에 제시된 기댓값은 집단 내 조우성과 만우성의 유전자 및 유전자 빈도(Table 1)를 근거로 계산된 값이다. 분석 결과, 모든 교배조합에서 관측치와 기대치 간의 유의적 차이가 없는 것으로 나타나, 이론적 분리비와 실제 교배 검정에 따른 분리비가 일치한다고 보여진다(P>0.05). 특히 병아리의 깃털 자가성감별을 위해서는 조우성 ♂ × 만우성 ♀의 교배조합이 조성되어야 하고, 이들로부터 생산된 수평아리는 모두 만우성이고, 암평아리는 모두 조우성이어야 한다. 시험 결과, 본 교배조합으로부터 생산된 총 140수의 병아리 중 수컷에서 2수의 오차가 발생하여 98.6%의 감별 일치율을 보였다. 따라서 한국재래닭 적갈색종을 이용하여 깃털 자가성감별 계통을 조성하였을 때 병아리의 깃털 조만성 형태로 암수 감별이 유효한 것으로 판단되어 이의 산업적 활용 가능성을 제시하는 바이다.