
INTRODUCTION
The worldwide consumption of chicken meat has been gradually increasing. According to a United Nations Food and Agriculture Organization (UN-FAO) report, the consumption of poultry meat is growing rapidly, and the annual per capita consumption worldwide is expected to reach 17.2 kg in 2030. This report predicts that poultry meat consumption will increase continuously because it is an inexpensive, good quality protein resource. Although chicken is the fowl eaten the most worldwide, turkey and duck are also important, mainly in the United States and China.
This increasing consumption trend reflects the nutritional superiority of poultry meat. In terms of fat composition, poultry meat contains less fat and cholesterol and fewer saturated fatty acids than other meat products. Moreover, the omega-3/omega-6 fatty acids ratio is higher in poultry meat than in other meat products. The fatty acid composition can affect meat quality traits, such as firmness, shelf life, and flavor. Specifically, the volatile, odorous, and lipid oxidation process during cooking can affect the odor and flavor of meat (Wood et al., 2004). Therefore, the fatty acid composition is an important parameter when evaluating the quality and taste of meat.
Chicken meat has the largest market share in the Korean poultry industry. The demand for native chicken meat is increasing steadily due to an increase in consumer preference for high-quality meat. Korea has five distinct Korean native chicken (KNC) lines that are adapted to the regional climate and resistant to disease. Additionally, KNC meat has a high protein content and a unique texture and flavor in comparison with commercial broilers. The excellent flavor of KNC is attributable to the fact that it is rich in C18:2, C20:4, C20:5, and C22:6 fatty acids, which are well-known taste-active components of chicken meat (Jin et al., 2017).
The fatty acid composition is affected by various environmental factors, and it has difficulties as to what criteria should be used selecting the birds for the actual breeding program. In particular, the interactions of genes are important, as various genes affect fatty acid composition in meat. Through the previous study, we investigated the relationships between the fatty acid composition and nine single-nucleotide polymorphisms (SNPs) of six candidate genes (DEGS1, ELOVL6, FABP3, FABP4, FASN and SCD) in KNC. We found total 10 significant SNP effects (P<0.05), 21 significant haplotype effects (P<0.05) and 51 significant SNP x SNP pairwise interactions (P≤0.05) in thigh and breast meat (Jin et al., 2020). Therefore, in this study, we performed additional analysis of the patterns of SNP × SNP interactions which obtained through previous studies. We investigated the epistatic effects from the combination of two genotypes affecting one fatty acid trait in order to utilize in the future breeding programs. The results of this study could be used as information to improve the meat quality, especially fatty acid composition, of KNC.
MATERIALS AND METHODS
This study followed “The Guide for Care and Use of Laboratory Animals” published by the Institutional Animal Care and Use Committee of the National Institute of Animal Science (NIAS) in Korea. The detailed information on the materials and methods used in this study was already described in Jin et al. (2020). Briefly, the thigh and breast meat from 597 KNC were used for this study, the lipid samples were extracted using Folch method (Folch et al., 1957) and the fatty acid composition (%) was analyzed using gas chromatography (HP 7890, Agilent Technologies, USA). Five SNPs of four candidate genes (FABP3, FABP4, FASN and SCD) were genotyped using PCR-restriction fragment length polymorphism (PCR-RFLP) and four SNPs in two genes (DEGS1 and ELOVL6) were genotyped using the BioMark Fluidigm array system (Fluidigm® 192.24 SNP Type™, USA). The genotype information is represented at the Table 1 The effects of SNP × SNP interactions were investigated using linear mixed model in the ASReml-W software (VSN International, Hemel Hempstead, UK).
where Yijklm is the observation for trait, μ is the overall mean, Li is the fixed effect of the ith line (five levels), Sj is the fixed effect of the jth sex (two levels), Bk is the fixed effect of the kth batch (two levels), and Gl is a SNP combination coded as 0, 1 and 2 as a covariate of each genes. CWm is the covariate for the carcass weight, Aijklm is a vector of random polygenic effect) and eijklm is a vector of random residual . Among them, of the 50 results with P values less than 0.05 were used for this study. Python and R programs were used for illustrating the figures of SNP × SNP interaction patterns.
RESULTS AND DISCUSSION
The significant epistatic effects were found in all fatty acids except for C20:1 in thigh and C16:0 in breast meat. The patterns of genotype combination which influenced the phenotype were appeared very diverse. As the genotypic value coded as 0, 1, and 2 were increased (substituted to alternative allele), the phenotypic values were changed as follows, 1) increased or decreased together, 2) when the genotypic value is 1 (the allele is heterozygote), the phenotypic value is the highest or lowest, and then the genotypic value is 2, the phenotype value is increased or decreased compare to that of genotype value is 0. Following these patterns, the genotype combination can contribute to raising or lowering the phenotype value (meat fatty acid composition, %). In the case of the epistatic effect on C18:3 in the thigh meat, when the genotype value of rs313838448 in SCD and rs316253049 in FABP4 changed from 0 to 1, both the phenotype values showed a tendency to decrease. On the other hand, in the case of rs313838448 in SCD and rs741339687 in DEGS1, the phenotype value of rs313838448 in SCD decreased, whereas the phenotype value of rs741339687 in DEGS1 increased, and the genotype value showed a big difference in phenotype value at 2. As the analysis results of this study on C20:4, five epistatic effects were identified for each thigh and breast meat. Among them, four combination effects (rs313838448 in SCD and rs316253049 in FABP4, rs313838448 in SCD and rs741339687 in DEGS1, rs10731498 in SCD and rs741339687 in DEGS1, rs316253049 in FABP4 and rs313000917 in ELOVL6) appeared in both parts of meat, but the patterns were different (Fig. 1, 2).
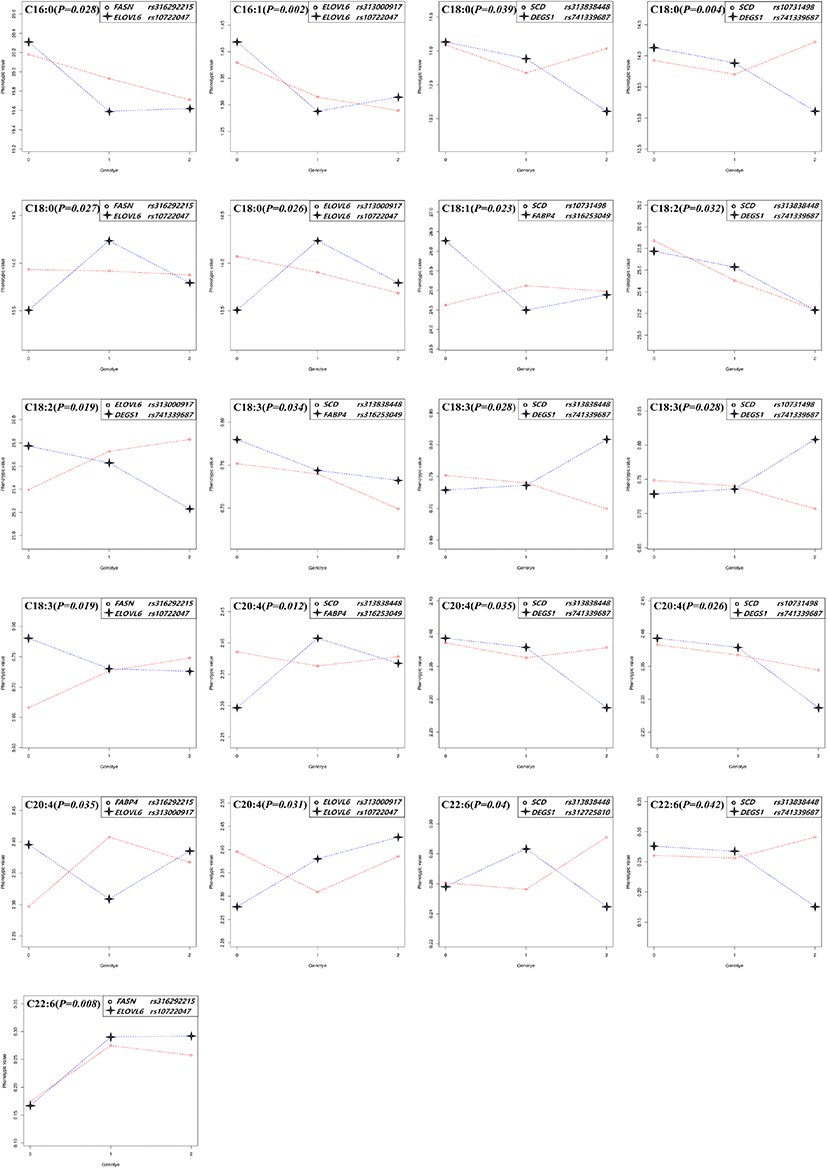
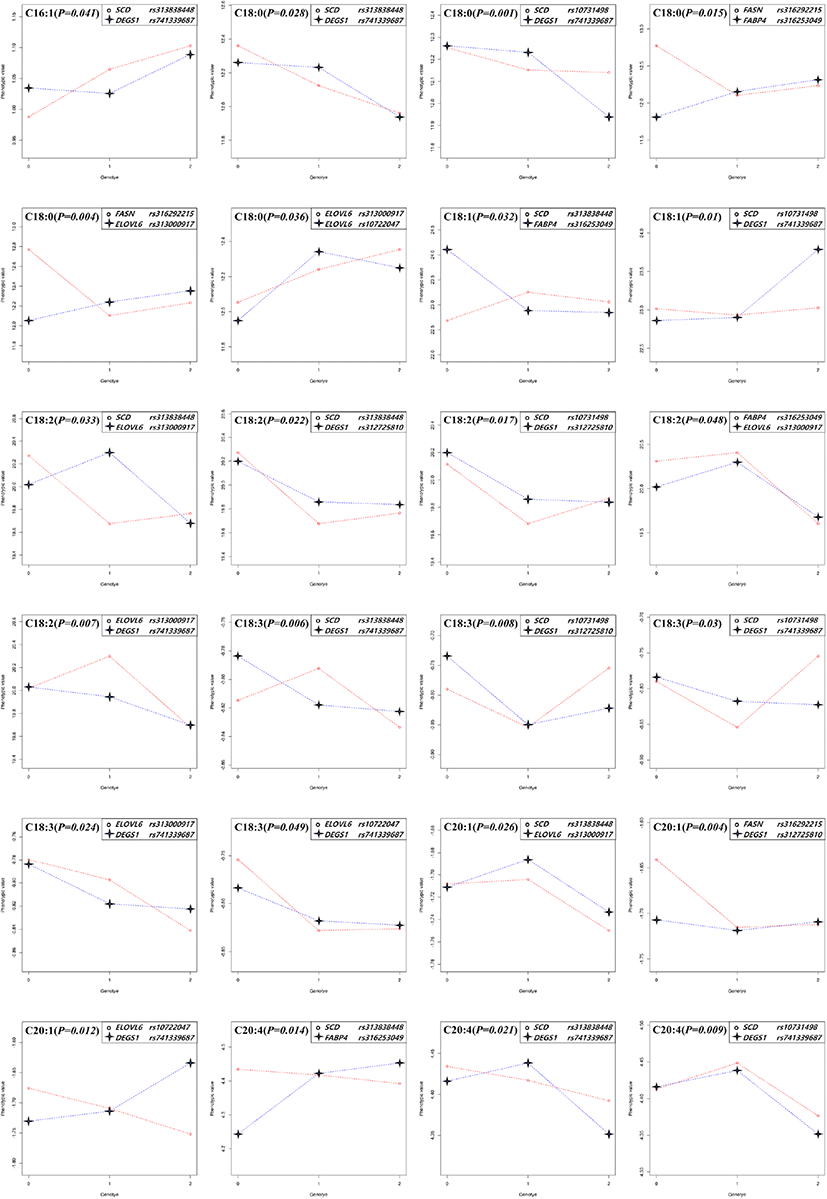
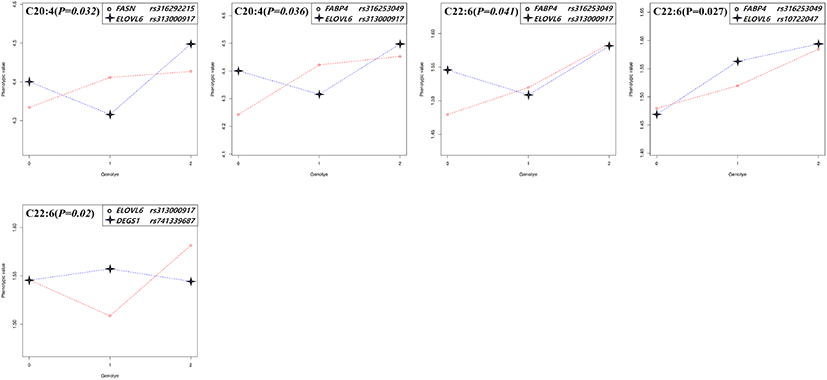
Genetic relationship studies of the fatty acid composition of meat mainly have examined in beef and pork. In these, the SCD and FASN genes were related to the meat fatty acid composition, and they contributed to increased monounsaturated fatty acids (MUFAs), especially in beef (Taniguchi et al., 2004; Zhang et al., 2008). Moreover, various studies have reported that both candidate genes had significant genetic relationships with the C18:1 composition of beef (Uemoto et al., 2011; Li et al., 2012; Yokota et al., 2012). Ishii et al. (2013) also reported that bovine SCD and FASN genes were related to the fatty acid composition, but they confirmed a relationship only for C14:1. For pork, SCD and FASN had significant effects on the fatty acid composition (Renaville et al., 2013; Grzes et al., 2016; Henriquez-Rodriguez et al., 2016; Sato et al., 2017). Uemoto et al. (2011) reported a strong effect of SCD on the fatty acid composition and fat melting point using quantitative trait locus (QTL) fine mapping. The main role of FABP genes is to regulate fatty acid uptake and transport, and they are specifically distributed in tissues where they perform their respective roles.
FABP3 (heart FABP) and FABP4 (adipocyte FABP) have been widely reported to affect the fats in beef and pork. Lee et al. (2010) also reported that FABP3 affects the unsaturated fatty acid (UFA) content of pork. Oh et al. (2012) reported a significant effect of FABP4 on UFAs, whereas Narukami et al. (2011) and Hoashi et al. (2008) reported its effects on C16:0 and C16:1, respectively, in beef. ELOVL6 is also involved in fatty acid synthesis, and Corominas et al. (2013) reported that porcine ELOVL6 was strongly associated with C16:0 and C16:1 in muscle and backfat. The DEGS1 gene is an endoplasmic reticulum membrane enzyme that catalyzes the desaturation of fatty acids in humans (Cadena et al., 1997), although there are no specific reports on its effects on the fatty acid composition of meat.
Compared with studies of beef and pork, relatively few chicken meat studies have examined the genetic relationships of candidate genes with the fatty acid composition. Maharani et al. (2011; 2013) investigated SCD, FASN, FABP3, and FABP4 in broiler chicken meat and found that the SCD gene affected C14:0, C16:0, and C16:1, whereas the other genes had no significant effects. D’Andre et al. (2013) reported the effects of FASN and ELOVL6 on fat deposition by comparing fast- and slow-growing chickens. Wang et al. (2006) and Wang et al. (2007) also reported the effects of FABP4 on lipid deposition and various fatness traits. Fatty acid composition and fat deposition are closely related to each other. Fat deposition is determined by de novo fatty acid synthesis and the uptake of exogenous fatty acids (De Smet et al., 2004). Fat deposition is a major factor that affects the fatty acid composition of pig and ruminant meats (Wood et al., 2004).
Considering these SNP effects on meat fatty acid composition, it is necessary to investigate the epistatic effects of SNP genotype combinations. The epistatic effect is the interaction between multiple genetic variants located at different genetic loci. This means that the phenotypic effect of having more than one genetic variation may be different from the effect of simply adding the effects of each variation(Lee et al., 2006). These effects have been investigated for complex traits in human diseases (Cordell, 2002), and polygenic traits in livestock. Regarding the fatty acid composition of beef, Matsuhashi et al. (2011) identified the epistatic effects of five SNPs of four genes, FASN, SCD, SREGP1 and GH, in longissimus muscle of Japanese black cattle and reported the significance between GH and SCD on C18:3. Li et al. (2012) investigated the interaction effects between SCD and FASN in brisket adipose tissue of Canadian Angus and Charolais-based commercial cross bred, and identified significant epistatic effects on various fatty acids. Therefore, we need to consider the difference in genotype effect according to allele substitution in the improvement of polygenic traits such as fatty acids. In addition, the epistatic effect on the phenotype should checked to predict the transmission patterns to the next generation, and non-additive effects should be researched from various areas.
The fatty acid composition of meat is influenced more by environmental factors than by genetic factors (De Smet et al., 2004; Wood et al., 2004). Nonetheless, many genetic studies have examined genetic variations based on heritable characteristics (Ishii et al., 2013). In this aspect, it is necessary to investigate the differences between varieties or lines based on the genetic diversity of chickens, and it is thought that genetic parameters are useful in the livestock industry. It is necessary to determine the fatty acid composition preferred by consumers and understand the synthesis and metabolism of fatty acids.
This study provides genetic information on the fatty acid composition of chicken meat. In particular, the results of this study should be useful for improving KNC, which has unique flavors and characteristics compared with commercial broilers.