
서 론
가금류 생산에서 환경 스트레스는 생산성, 건강 및 육질에 중대한 영향을 미치는 핵심 요인으로 인식되어 왔다. 최근 기후변화로 인하여 폭염, 한파 및 이상 기온 등의 영향이 급속도로 커지고 있으며, 사계절이 뚜렷한 한국의 경우 기온이 동절기에는 최저 −30°C까지 떨어지고 하절기에는 최대 40°C까지 상승할 수 있다(Seo et al., 2009). 육계가 저온에 노출될 경우 육계는 체온 유지를 위해 추가 에너지를 소비하며, 이는 사료 효율의 감소와 생산성 저하로 이어질 수 있다(Park, 2008). 한국에서는 겨울철 가금류 생산에서 적절한 사육 환경을 유지하는 것이 주요한 과제이며, 생산성을 유지하고 저온 노출을 방지하기 위해 난방이 필수적이다. 이 중 저온 스트레스(cold stress)는 겨울철 사육 환경이나 환기 관리가 미흡한 축사 조건에서 빈번하게 발생하며, 특히 병아리와 육계에서 성장 저하, 사료효율 악화 및 면역 기능 저하를 유발하는 주요 환경적 도전 요인으로 작용한다(Su et al., 2018; Xue et al., 2021). 저온 스트레스는 육계의 경우, 주변 온도가10°C 이상 급격히 감소하거나 또는 주변 온도보다 4°C 이상 낮은 온도에 장시간 노출될 때, 또는 최적 열중성대(thermal neutral zone)보다 10°C 이상 낮은 환경에 노출되는 것을 의미하며(Liu et al., 2022), 겨울철 환경 온도가 18°C 이하로 떨어지는 경우를 저온 스트레스로 간주하기도 한다(Kim et al., 2023). 일반적으로 성계 가금류의 최적 열중성대는 약 18~24°C로 알려져 있으며(Kim et al., 2023), 이 범위를 벗어난 저온 환경에서는 체온 유지를 위한 대사적 부담이 급격히 증가한다. 가금류는 부화 초기에는 체온 조절 능력이 제한적이며, 성장에 따라 점차 항온동물로서의 체온 항상성을 확립한다(Gonzalez-Rivas et al., 2020; Moreira et al., 2024). 특히 어린 병아리는 저온 환경에 취약하며, 성계에서도 주변 온도가 16°C 이하로 낮아질 경우 저온 스트레스로 인한 생리적·대사적 변화가 유발될 수 있다(Kim et al., 2023). 이러한 저온 스트레스는 산란계의 생산성과 난질을 저하시킬 뿐만 아니라(Li et al., 2020; Rajabi and Torki, 2021), 육계의 성장 성적 저하 및 복수수종 발생 위험 증가와도 연관되는 것으로 보고되었다(Ipek and Sahan, 2006; Wideman, 2001).
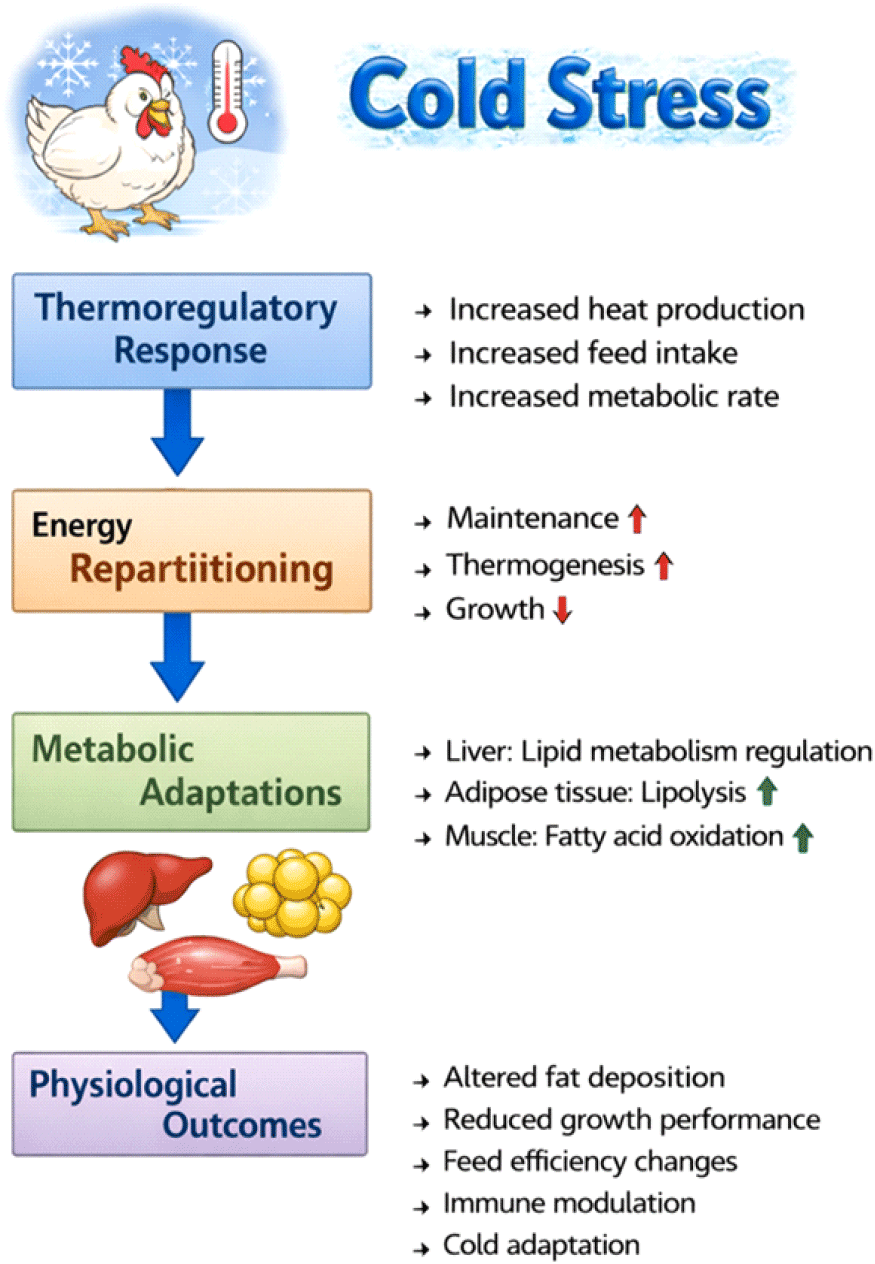
저온 노출 시 가금류는 체온 유지를 위해 대사율을 증가시키며, 이 과정에서 유지 에너지 요구량이 증가하고 체내 에너지 이용 전략이 근본적으로 재편된다(Gong et al., 2023; Li et al., 2024). 최근 연구들은 저온 스트레스가 단순한 체온 저하 문제가 아니라, 에너지 분배(energy distribution)를 재조정하는 대사적 스트레스 요인임을 강조하고 있다(Gong et al., 2023). 그러나 기존 환경 스트레스 연구는 주로 열 스트레스에 집중되어 왔으며(Nawab et al., 2018; Balakrishnan et al., 2023), 저온 스트레스에 따른 대사적 적응 기전에 대한 체계적인 논의는 상대적으로 부족한 실정이다. 최근 들어 저온 스트레스가 미토콘드리아 활성 증가와 지질대사 재편을 동반하며, 이 과정에서 활성산소종(ROS) 생성과 산화스트레스가 유발된다는 점이 보고되고 있다(Zhao et al., 2014; Ouchi et al., 2021). 이에 대응하여 항산화 방어 시스템과 heat shock proteins(HSPs)의 발현이 증가하며, 이러한 세포 보호 기전은 저온 환경에서의 대사 항상성 유지에 중요한 역할을 수행하는 것으로 제시되고 있다(Balakrishnan et al., 2023). 특히 저온 스트레스하에서의 지질대사 변화는 열 생산과 에너지 항상성 유지를 위한 핵심 적응 기전으로 주목받고 있다(Gong et al., 2023; Ahmadipour et al., 2025). 닭의 저온 스트레스 영향에 대한 개념적 개요는 Fig. 1에 제시하였다.
이에 본 총설은 가금류에서 저온 스트레스가 유발하는 에너지 분배 재프로그래밍과 지질대사 변화를 중심으로, 산화스트레스 및 HSPs의 역할을 통합적으로 고찰하고자 한다. 또한 저온 스트레스로 인한 부정적 영향을 완화하기 위한 영양적·관리적 전략에 관한 최신 연구 동향을 정리함으로써, 향후 가금류 생산 현장에서 활용 가능한 생리·대사 기반 대응 전략 수립에 기초 자료를 제공하고자 한다.
본 론
저온 스트레스는 가금류에서 단순한 환경적 불편 요인이 아니라, 전신적인 대사 재편(metabolic reprogramming)을 유도하는 강력한 생리적 자극으로 작용한다. 저온 환경에 노출되면 가금류는 체온 항상성을 유지하기 위해 기초 대사율을 증가시키고, 이로 인해 에너지 요구량이 급격히 상승한다. 이러한 반응은 특히 체온 조절 능력이 제한적인 어린 병아리나 고속 성장 육계에서 두드러지며, 에너지 섭취와 이용의 균형이 쉽게 붕괴될 수 있다(Gong et al., 2023; Li et al., 2024). 병아리(6일령 이전)가 저온 스트레스에 노출되면 생체 대사에 수주간 영향을 미칠 수 있으며 복수수종(ascites)도 증가한다(Malan et al., 2003). 저온 스트레스하에서 가장 두드러진 변화 중 하나는 에너지 사용의 우선순위가 성장과 생산에서 체온 유지와 생존으로 전환된다는 점이다. 저온 자극은 유지 에너지(maintenance energy) 요구량을 증가시키고, 상대적으로 성장, 지방 축적, 혹은 생산 관련 대사 경로로 배분되던 에너지를 열 생산(thermogenesis)에 집중시키는 방향으로 재조정한다. Gong et al.(2023)은 육계에서 적절한 cold stimulation(저온 자극)이 에너지 분배 양상을 변화시켜 스트레스 저항성을 향상시킨다고 보고하였으며, 이는 저온 스트레스가 수동적인 손상이 아니라 능동적인 대사 적응 반응임을 시사한다. 이러한 대사 적응의 중심에는 미토콘드리아 기능 변화와 열 생산 증가가 자리하고 있다. 저온 노출은 미토콘드리아의 산화적 인산화 활성을 촉진하여 ATP 생성뿐 아니라 열 발생을 증가시키며, avian uncoupling protein(av-UCP)과 같은 분자의 유전적 변이 또한 저온 노출 시 열 생산 효율에 영향을 미치는 것으로 보고되었다(Ouchi et al., 2021). 이 과정에서 에너지 효율은 감소하지만, 체온 유지를 위한 생존 전략으로서 의미를 갖는다.
한편, 저온 스트레스에 대한 대사 반응은 노출 방식과 강도에 따라 상이한 결과를 나타낼 수 있다. 육계에서 지속적인 저온 스트레스는 성장 저하와 산화 손상을 유발하는 반면, 간헐적 또는 조절된 저온 자극은 심장과 같은 주요 대사 기관에서 에너지 대사를 개선하고 스트레스 적응 능력을 향상시킬 수 있음이 보고되었다(Li et al., 2024). 이는 저온 스트레스가 항상 부정적인 자극만은 아니며, 조건에 따라 대사적 가소성(metabolic plasticity)을 유도하는 자극으로 작용할 수 있음을 의미한다. 그러나 과도하거나 지속적인 저온 스트레스는 대사 적응 범위를 초과하여 심장 조직에서 산화스트레스, 염증 반응 및 세포 사멸 경로를 동시에 활성화할 수 있음이 보고되었다(Wei et al., 2023).
종합하면, 저온 스트레스는 가금류에서 체온 저하에 따른 수동적 반응이 아니라, 에너지 요구 증가, 대사 경로 재조정, 그리고 기관․조직 수준의 기능 변화를 동반하는 복합적 대사 도전 요인으로 이해되어야 한다. 이러한 관점은 저온 스트레스하에서 관찰되는 지질대사 변화, 산화스트레스 증가, 그리고 세포 보호 기전 활성화에 대한 통합적 이해의 출발점이 되며, 다음 절에서 다룰 에너지 분배 재편과 지질대사 조절 기전을 해석하는 핵심 개념적 틀을 제공한다.
저온 스트레스에 노출된 가금류에서 가장 근본적인 생리적 변화는 유지 에너지 요구량(maintenance energy requirement)의 증가이다. 저온 환경에서는 체온 항상성 유지를 위한 열 손실이 증가하며, 이를 보상하기 위해 기초 대사율이 상승한다. 그 결과, 동일한 사료 섭취 조건에서도 성장이나 생산에 이용될 수 있는 가용 에너지는 감소하고, 에너지 분배의 우선순위가 유지 및 생존 중심으로 재편된다. 실제로 육계에서 저온 스트레스는 사료효율 저하와 성장률 감소를 동반하며, 이는 증가된 유지 에너지 요구량이 주요 원인으로 지적되고 있다(Zhou et al., 2021; Gong et al., 2023; Li et al., 2024). 유지 에너지 요구 증가에 대응하는 핵심 대사 반응은 열 생산(thermogenesis)의 활성화이다. 가금류는 포유류와 달리 전형적인 갈색지방조직을 갖지 않지만, 미토콘드리아 기능 조절을 통해 효율적으로 열을 생성한다. 저온 스트레스하에서는 미토콘드리아의 산화적 대사가 항진되고, ATP 합성과 더불어 열 발생이 증가하여 체온 유지에 기여한다. 이러한 열 생산 증가는 에너지 효율의 저하를 동반하지만, 저온 환경에서의 생존을 위한 필수적인 대사 전략으로 간주된다(Gong et al., 2023). 열 생산 과정에서 중요한 특징은 에너지 이용 효율의 의도적 감소이다. 미토콘드리아에서 산화적 인산화가 부분적으로 비결합(uncoupling)되면서, 에너지가 성장이나 조직 축적보다는 열 형태로 분산된다. 이로 인해 지방산 산화와 같은 에너지 공급 경로의 중요성이 증가하며, 지질대사가 저온 스트레스 적응의 핵심 축으로 부상하게 된다. 이러한 에너지 분배의 변화는 이후 지질대사 재편과 산화스트레스 증가로 직결된다. 조류에서는 하나의 UCP(av-UCP)가 발견되었으며, 이는 포유류의 UCP2 및 UCP3와 높은 상동성을 가진다(Evock-Clover et al., 2002; Toyomizu et al., 2002). Av-UCP는 호흡의 비결합(respiration uncoupling) 수준에 영향을 미칠 수 있으며, 미토콘드리아 내막을 가로질러 이동하는 지방산을 조절하는 역할도 한다(Richards, 2003). 이와 관련하여, Av-UCP은 저온 스트레스하 에너지 분배와 열 생산을 조절하는 중요한 유전적 요인으로 주목받고 있다. Ouchi et al.(2021)은 Av-UCP의 단일염기다형성(single nucleotide polymorphism)이 병아리의 저온 노출 시 열 생산량에 영향을 미친다고 보고하였으며, 이는 유전적 배경에 따라 저온 스트레스에 대한 대사 적응 능력이 달라질 수 있음을 시사한다. Av-UCP의 활성 증가는 미토콘드리아 내 양성자 누출을 촉진하여 열 발생을 증가시키는 반면, ATP 생성 효율을 감소시켜 에너지 분배 양상에 직접적인 영향을 미친다(Ouchi et al., 2021). 저온 스트레스는 온도에 의한 자극이라는 점에서 열적 스트레스에 해당하지만, 고밀도 사육과 같은 비열적 환경 스트레스 요인들 또한 가금류에서 유사하게 대사 재프로그래밍을 유도하며, 이는 열충격단백질(HSPs), 소포체 스트레스 지표(ER stress markers), 그리고 지질 및 포도당 대사 관련 유전자들의 발현 변화로 특징지어진다(Sohn et al., 2015). 열 스트레스는 닭에서 간의 지질대사를 체계적으로 재편하는 것으로 보고되었으며, 이는 de novo 지방산 생합성의 동시적 증가, 지방산 산화의 억제, 그리고 VLDL 분비 장애로 특징지어진다. 이러한 개념적 틀은 저온 스트레스를 포함한 다른 환경 스트레스 요인들이 가금류에서 에너지 분배 양상을 어떻게 재편하는지를 이해하는 데 중요한 기준을 제공한다(Moon, 2025).
종합적으로, 저온 스트레스는 가금류에서 유지 에너지 요구 증가를 출발점으로 열 생산을 강화하고, 유전적 요인에 의해 조절되는 미토콘드리아 기능 변화를 통해 에너지 분배를 재편하는 과정으로 이해될 수 있다. 이러한 에너지 분배의 재편은 단순한 대사 비용 증가를 넘어, 지질대사 조절, 산화스트레스 유발, 그리고 세포 보호 기전 활성화로 연쇄적으로 확장된다.
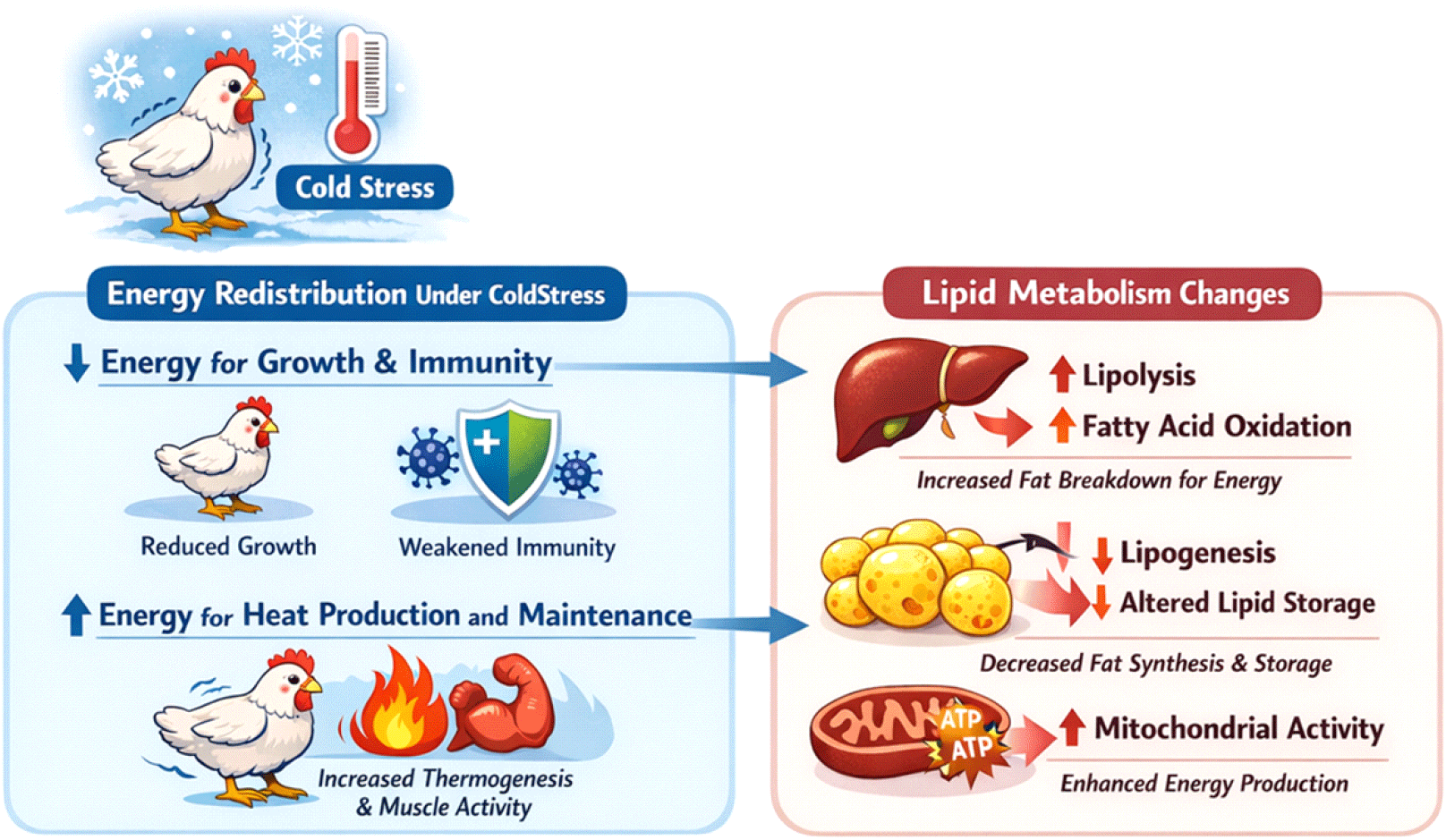
저온 스트레스로 인한 에너지 분배의 재편은 필연적으로 지질대사의 구조적, 기능적 변화를 동반한다. 유지 에너지 요구 증가와 열 생산 활성화로 인해, 지질은 저온 스트레스하에서 가장 중요한 에너지원으로 활용되며, 이 과정은 간, 지방조직, 근육에서 조직 특이적 방식으로 조절된다. 이러한 지질대사 재편은 단순한 지방 축적 변화에 그치지 않고, 산화스트레스, 육질 및 생산성에 장기적인 영향을 미친다(Gong et al., 2023).
지질은 고에너지 유기분자로서, 합성과 분해 과정을 조절함으로써 체온 조절에 중요한 역할을 한다. 조류에서 간은 지질대사와 에너지대사를 조절하는 주요 기관이다(Nguyen et al., 2008). 간은 지방산 합성의 중요한 장소이며(Ramachandran et al., 2007), 이후 발생하는 대사적 열 생산은 체온 유지와 저온 스트레스에 대한 대응에 핵심적인 역할을 한다(Sepa-Kishi et al., 2018). 가금류에서 간의 지방산 생합성이 지질 합성의 주요 경로임을 고려할 때, 저온 스트레스로 인한 에너지 요구량 증가는 필연적으로 간에 대사적 부담을 가하게 되며, 그 결과 지질대사 흐름과 에너지 분배 양상에 중대한 변화를 초래한다(Moon, 2018). 간은 가금류에서 지질 합성과 수송의 중심 기관으로, 저온 스트레스하에서 가장 민감한 대사 변화를 보인다. Grefhorst et al.(2018)은 동물을 4°C 환경에 24시간 노출시켰을 때 간 내 중성지방과 담즙산 함량은 증가하였으나, 콜레스테롤 수준에는 영향을 미치지 않았다고 보고하였으며, 이는 저온 환경이 간의 지질대사 과정에 변화를 유도할 수 있음을 시사한다. 그러나 유사한 대사 기전이 포유류에서 보고된 바 있으나, 비교생리학적 종의 차이로 인해 이를 조류에 직접 적용하는 데에는 주의가 필요하다. 이와 같이 저온 환경에서는 열 생산을 위한 에너지 공급 요구가 증가함에 따라, 간에서의 지방산 동원과 대사 조절이 재편된다. 일부 연구에서는 저온 스트레스가 간 내 지방 합성 관련 유전자 발현을 변화시키는 동시에, 지방산 산화를 촉진하거나 지질 수출(VLDL 조립 및 분비)을 조절한다고 보고하였다(Zhang et al., 2024b). 이러한 변화는 에너지 항상성 유지에 기여하지만, 장기적인 저온 스트레스 상황에서는 간 내 지질 축적과 대사 부담을 증가시킬 수 있다(Ahmadipour et al., 2025). 특히, 저온 스트레스하에서는 미토콘드리아 활성 증가로 인해 간 내 산화적 대사가 항진되며, 이는 지질대사 재편과 산화스트레스 증가를 동시에 유발할 수 있다. 간에서의 이러한 대사적 적응은 이후 전신 지질 이용 효율과 직결되며, 저온 스트레스에 대한 전반적인 대사 적응의 핵심 요소로 작용한다.
지방조직은 저온 스트레스하에서 에너지 저장 기관에서 에너지 공급 기관으로 기능적 전환을 겪는다. 저온 자극은 지방 분해(lipolysis)를 촉진하여 자유 지방산의 방출을 증가시키며, 이는 열 생성과 유지 에너지 요구를 충족시키는 주요 기질로 활용된다. 최근 연구에 따르면, 저온 스트레스는 단순히 지방량을 감소시키는 것을 넘어, 지방조직의 구조적 재편과 세포 간 상호작용 변화를 유도한다. Zhang et al.(2024a)은 저온 스트레스 조건에서 병아리 피하지방조직에서 지방세포와 대식세포 간 상호작용이 증가하며, 혈관형성과 조직 재구성이 촉진된다고 보고하였다. 이러한 변화는 지방조직의 대사적 유연성을 향상시키고, 지속적인 에너지 공급을 가능하게 하는 적응 반응으로 해석된다(Zhang et al., 2024a). 또한, 조류 피부 및 피하지방의 발달 가소성에 관한 연구는 저온 스트레스가 지방조직 발달 경로 자체를 재조정할 수 있다(Ou et al., 2025). 최근 연구들은 가금류에서 저온 스트레스에 의해 유도되는 지방조직 재편이 단순한 지방 분해 증가나 구조적 변화에 국한되지 않음을 보여준다. Mezentseva et al.(2008)은 닭 배아에서 유래한 중간엽 세포가 시험관 내(in vitro)에서 포유류 성숙 갈색지방세포의 형태학적·생화학적 특성을 지닌 갈색지방세포-유사 세포(brown adipocyte-like cells)로 분화할 수 있음을 보고하였다. 특히 Sotome et al.(2021)은 저온 환경에 노출된 닭(in vivo)에서 beige-like 지방세포의 출현을 최초로 입증하였으며, 이를 통해 조류에서도 유도성 열 발생 지방 조직(thermogenic adipose plasticity) 이 존재함을 제시하였다. 조류는 포유류와 달리 전형적인 갈색지방조직을 보유하지 않지만, 저온 노출 시 백색지방조직이 beige-like 상태로 표현형 전환되는 현상이 관찰되었다. 그러나 조류에서 beige-like 지방세포에 대한 현재의 근거는 아직 제한적이며, 그 기능적 중요성은 완전히 규명되지 않은 상태임을 유의해야 한다. 저온 스트레스하에서 beige-like 지방의 유도는 지방조직이 에너지 분배를 조절하는 능동적 조절자로 작용할 잠재적 가능성을 시사한다. 이는 저온 스트레스하에서 beige-like 지방의 유도는 지방조직이 에너지 분배를 조절하는 능동적 조절자로 작용함을 시사한다. 즉, 지방 저장 중심의 대사 상태에서 벗어나, 지질대사를 열 생산 방향으로 전환함으로써 체온 항상성 유지에 기여한다. 이러한 관점은 저온 스트레스가 조직 특이적인 에너지 대사 재프로그래밍을 유도하며, 이 과정에서 지방조직이 핵심적 역할을 수행한다는 개념을 뒷받침한다. 특히 이러한 지방조직의 가소성은 근육의 과도한 떨림성 열생성(shivering thermogenesis)이나 간 중심의 지방산 산화에 대한 의존도를 완화함으로써, 전신적인 대사 안정성 유지에 기여할 가능성이 있다. 가금에서 저온 스트레스에 의한 에너지 재분배 및 지질대사 재편에 대한 관계는 Fig. 2에 제시하였다.
닭고기의 품질은 다양한 잠재적인 내적 및 외적 요인의 영향을 받는다(Al-Sagan et al., 2021; Weimer et al., 2022). 온도 변화는 닭고기의 맛과 품질에 영향을 미치는 불가피한 외적 요인으로 여겨져 왔다. 지나치게 높은 온도 또는 낮은 온도는 열 스트레스나 저온 스트레스를 유발할 수 있으며, 이로 인해 닭고기의 맛과 품질이 영향을 받는 것으로 보고되었다(Henrikson et al., 2018; Maynard et al., 2023). 근육 조직은 저온 스트레스하에서 에너지 소비와 생산성의 균형을 동시에 반영하는 핵심 조직이다. 저온 환경에서는 근육에서의 지방산 이용이 증가하며, 이는 ATP 생성과 열 발생을 위한 중요한 에너지원으로 작용한다. 그러나 이러한 대사 변화는 근육 내 지질 조성, 미토콘드리아 기능, 그리고 단백질 구조에 영향을 미쳐 최종적으로 육질에 변화를 초래할 수 있다(Xu et al., 2021; Teng et al., 2022). 최근 연구에서는 저온 스트레스 조건에서 항산화 능력을 강화하거나 지질대사를 조절하는 영양적 개입이 근육 조직의 산화 손상을 완화하고 육질을 개선할 수 있음을 보고하였다. Zhang et al.(2026)은 루틴(rutin) 보충이 cold-induced oxidative stress를 감소시키고 근원섬유 단백질 구조를 개선하여 육질을 향상시킨다고 보고하였으며, 이는 근육 지질대사와 산화스트레스 간의 밀접한 연관성을 시사한다.
이상의 연구들을 종합하면, 저온 스트레스하에서의 지질대사 재편은 간의 대사 조절, 지방조직의 동원 및 구조적 적응, 그리고 근육의 지방산 이용 증가가 유기적으로 연결된 통합적 과정으로 이해될 수 있다. 이러한 조직 특이적 지질대사 변화는 열 생성과 에너지 항상성을 유지하는 데 필수적이지만, 동시에 미토콘드리아 ROS 생성 증가와 산화스트레스를 유발할 가능성을 내포한다. 특히 산란계에서는 저온 스트레스하에서 조단백질, 메티오닌 및 아연 수준을 조절한 식이 전략이 생산성과 난질을 개선할 수 있음이 보고되어, 지질대사 조절과 더불어 단백질 및 미량영양소 대사의 중요성도 함께 고려되어야 함을 시사한다(Mohebbifar et al., 2025).
저온 스트레스하에서 관찰되는 에너지 분배 변화와 지질대사 재편은 필연적으로 산화 스트레스(oxidative stress)와 redox imbalance를 동반한다. 저온 환경에 적응하기 위해 가금류는 미토콘드리아 활성을 증가시키고 열 생산을 강화하지만, 이러한 대사적 적응은 동시에 ROS의 과잉 생성을 초래한다. 따라서 저온 스트레스는 가금류에서 에너지 항상성 유지와 산화적 손상 위험이 공존하는 대사적 이중 부담으로 작용한다(Oke et al., 2024).
저온 노출은 thermogenesis 활성화를 통해 미토콘드리아의 산화적 인산화 속도를 증가시키며, 이 과정에서 전자전달계로부터의 전자 누출이 증가하여 ROS 생성이 촉진된다. 특히 지방산 산화가 주요 에너지원으로 활용되는 상황에서는 미토콘드리아 내 산화 환원 반응의 부담이 가중되어 ROS 축적 가능성이 더욱 높아진다. 이러한 미토콘드리아 유래 ROS 증가는 간, 근육, 면역 기관 등 다양한 조직에서 관찰되며, 저온 스트레스하 대사 적응의 불가피한 부산물로 인식되고 있다(Zhao et al., 2014; Oke et al., 2024). 최근 연구에서는 저온 스트레스가 Nrf2/HO-1 및 NF-κB 신호 경로를 조절함으로써 산화 스트레스, 염증 반응, 그리고 세포 사멸을 동시에 유도할 수 있음이 제시되었으며, 이는 단순한 ROS 증가를 넘어 분자 신호 수준의 병리적 반응을 포함함을 의미한다(Wei et al., 2023). Nrf2/HO-1 신호전달 경로는 산화적 자극에 매우 민감하게 반응하며, 대사효소 유전자의 기저 및 유도성 발현을 다면적으로 조절함으로써 산화 스트레스 조건하에서 다양한 장기와 조직에서 중요한 항산화 및 세포 보호 효과를 수행한다(Bardallo et al., 2022). 미토콘드리아 ROS는 저농도에서는 신호전달 분자로 작용할 수 있으나, 저온 스트레스와 같이 지속적․과도한 생성이 이루어질 경우 세포 내 산화 환원 균형을 붕괴시키고 단백질, 지질 및 핵산 손상을 유발한다. 이러한 redox imbalance는 이후 항산화 방어 시스템의 활성화를 촉발하는 주요 자극 요인으로 작용한다.
저온 스트레스에 의해 증가한 ROS에 대응하기 위해 가금류는 항산화 방어 시스템을 활성화한다. Superoxide dismutase(SOD), catalase(CAT), glutathione peroxidase(GPx)와 같은 항산화 효소는 ROS 제거에 핵심적인 역할을 수행하며, 저온 노출 시 이들의 발현 및 활성이 증가하는 것으로 보고되었다. Zhao et al.(2014)은 저온 스트레스에 노출된 닭의 면역 기관에서 항산화 효소 활성과 함께 HSPs 발현이 동시에 증가함을 보고하여, 산화 스트레스 대응이 다층적 보호 기전을 통해 이루어짐을 시사하였다. 그러나 항산화 방어 능력에는 생리적 한계가 존재하며, 저온 스트레스의 강도와 지속 기간이 증가할수록 항산화 시스템의 보상 능력은 점차 감소할 수 있다. 이로 인해 ROS 제거와 생성 간 균형이 무너질 경우, redox imbalance가 심화되어 조직 수준의 기능 손상으로 이어질 가능성이 높아진다.
저온 스트레스로 유발된 산화 스트레스는 조직 특이적 기능 손상으로 발현된다. 간에서는 지질과산화 증가와 미토콘드리아 기능 저하가 지질대사 조절 능력을 약화시킬 수 있으며, 이는 간 내 대사 부담을 가중시킨다. 근육 조직에서는 산화적 손상이 근원섬유 단백질 구조 변화를 유발하여 육질 저하로 이어질 수 있고, 장기적으로는 성장 성적에도 부정적인 영향을 미칠 수 있다(Zhang et al., 2026). 또한, 면역 기관에서의 산화스트레스 증가는 면역 세포 기능 저하와 스트레스 감수성 증가로 연결되어, 저온 스트레스하에서 가금류의 질병 취약성을 높이는 요인으로 작용한다. 이러한 조직 기능 손상은 단순한 산화적 손상에 국한되지 않고, 에너지 대사 이상, 염증 반응, 세포 사멸 경로 활성화 등으로 확장될 수 있다(Zhao et al., 2014; Oke et al., 2024).
종합적으로, 저온 스트레스하에서의 산화스트레스는 미토콘드리아 ROS 과잉 생성, 항산화 방어 활성화, 방어 한계 초과 시 조직 기능 손상이라는 연속적 경로로 이해될 수 있다. 이러한 상황에서 세포는 항산화 효소 외에도 추가적인 보호 기전을 필요로 하며, 그 대표적인 예가 HSPs이다.
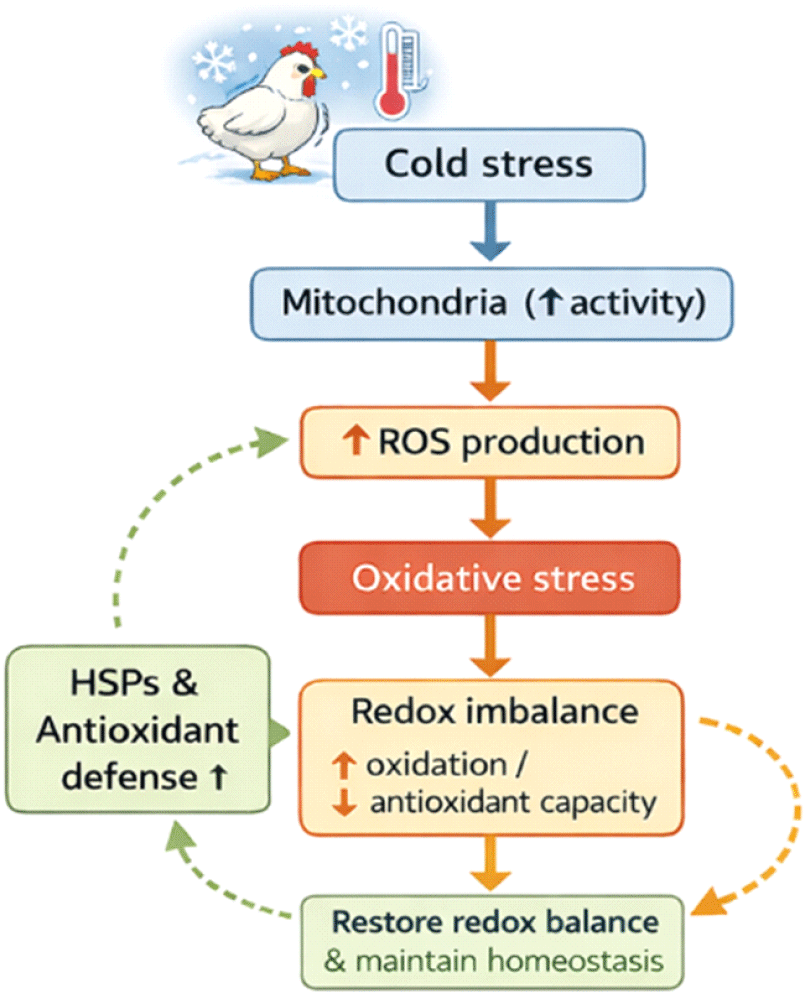
저온 노출은 신경내분비계, 항산화 시스템, 그리고 면역계의 기능에 영향을 미칠 수 있다(Onderci et al., 2003; Hangalapura et al., 2006). 더 나아가, 저온 스트레스가 면역 반응에 미치는 영향은 스트레스의 지속 시간과 강도에 따라 달라질 수 있다는 점도 제시되었다(Hangalapura et al., 2003). 또한 여러 연구에서 저온 노출이 항산화 방어 시스템에 영향을 미친다는 사실이 보고되었다(Lin et al., 2004; Mujahid, 2009). 생물체가 다양한 스트레스 조건에 노출되면 대부분의 단백질 합성은 억제되지만, 고도로 보존된 단백질 군 HSPs는 신속하게 합성되며, HSPs는 스트레스 반응을 조절하는 데 핵심적인 구성 요소이다(Al-Aqil and Zulkifli, 2009). HSP의 발현 수준은 정상적인 생리 조건에서는 낮지만, 저온 및 열 스트레스하에서는 스트레스 저항성을 향상시키기 위해 빠르게 증가할 수 있다(Wu, 1995; Wei et al., 2018). 따라서, 순환계 내에 HSPs가 증가하는 것은 숙주에게 위험 신호(danger signal)로 작용한다. 또한 Zhao et al.(2014)은 12±1°C의 저온 스트레스 조건에서 육계의 면역 기관에서 HSP27, HSP40, 그리고 HSP60의 mRNA 발현이 증가하였음을 보고하였다. 저온 스트레스로 유발되는 산화스트레스와 redox imbalance는 세포 수준에서 추가적인 보호 기전의 활성화를 요구한다. 이러한 상황에서 HSPs는 단백질 항상성 유지와 세포 생존을 보장하는 핵심 방어 인자로 작용한다. 비록 HSPs가 주로 고온 스트레스와 연관되어 연구되어 왔으나, 최근 연구들은 저온 스트레스 또한 가금류에서 HSP 발현을 유도하는 강력한 자극임을 명확히 보여주고 있다(Zhao et al., 2014; Moon, 2020; Balakrishnan et al., 2023).
저온 노출은 미토콘드리아 ROS 증가와 단백질 변성 위험을 동반하며, 이는 HSP 발현을 유도하는 주요 신호로 작용한다. 저온 스트레스 조건에서 산화 스트레스, HSP 활성화, 그리고 미토콘드리아 보호 간의 상호작용에 대한 제안된 기전은 Fig. 2에 제시되어 있다. Zhao et al.(2014)은 저온 스트레스에 노출된 닭의 면역 기관에서 HSP70을 포함한 여러 HSPs의 발현이 유의적으로 증가함을 보고하였으며, 이는 저온 환경에서도 HSPs가 세포 보호에 적극적으로 관여함을 시사한다. 이러한 HSP 발현은 단백질 접힘(protein folding) 보조, 변성 단백질의 재접힘 또는 분해 촉진을 통해 세포 손상을 최소화하는 방향으로 작동한다(Zhao et al., 2014). 저온 스트레스하에서의 HSP 발현 증가는 조직 특이적으로 나타날 수 있으며, 특히 대사 활성이 높은 간, 근육, 면역 기관에서 두드러진다. 이는 저온 스트레스에 대한 HSP 반응이 단순한 일반 스트레스 반응이 아니라, 대사 부담 증가에 대한 적응적 반응임을 시사한다.
HSPs는 직접적인 항산화 효소는 아니지만, ROS로 인한 세포 손상을 완충(buffering)하는 간접적 보호 역할을 수행한다. HSP70과 HSP90은 미토콘드리아 단백질과 항산화 효소의 구조적 안정성을 유지하고, 산화적으로 손상된 단백질의 축적을 억제함으로써 redox imbalance의 악화를 방지한다. Balakrishnan et al.(2023)은 가금류에서 HSPs가 산화스트레스 조건하에서 세포 생존 신호를 강화하고, 스트레스 적응 능력을 향상시키는 중심적 역할을 수행한다고 정리하였다. 또한, HSPs는 ROS에 의해 활성화되는 세포 사멸 경로를 억제하고, 스트레스 관련 신호전달 경로와 상호작용함으로써 세포 손상을 제한한다. 이러한 특성은 저온 스트레스하에서 항산화 시스템의 방어 능력이 한계에 도달했을 때, HSPs가 '제2의 방어선(second line of defense)'으로 기능함을 의미한다.
저온 스트레스에 의해 재편된 에너지 분배와 지질대사는 대사 효율 저하와 조직 기능 불안정을 초래할 수 있으며, 이 과정에서 HSPs는 대사 안정화(metabolic stabilization)에 기여한다. HSPs는 미토콘드리아 기능 유지와 효소 단백질의 안정성을 보조함으로써, 과도한 대사 변동으로 인한 기능 붕괴를 완화한다. 이는 열 생성과 지방산 산화가 지속적으로 요구되는 저온 스트레스 환경에서 특히 중요하다. HSPs의 이러한 보호 작용은 단기적으로는 세포 생존을 보장하고, 장기적으로는 조직 기능 유지와 생산성 저하 최소화로 이어질 수 있다. 결과적으로 HSP 반응은 저온 스트레스에 대한 단순한 방어 반응을 넘어, 에너지 대사, 산화 스트레스, 세포 항상성을 연결하는 통합 조절 축으로 이해될 수 있다(Balakrishnan et al., 2023).
종합하면, 저온 스트레스하에서의 HSP 반응은 ROS 증가에 의해 유도되고, 산화 손상을 완충하며, 재편된 대사 환경을 안정화하는 다기능적 보호 기전으로 작용한다. 이러한 HSP 중심의 세포 보호 전략은 항산화 효소 시스템과 상호 보완적으로 작동하여, 저온 스트레스로 인한 조직 손상과 생산성 저하를 완화한다. 가금에서 저온 스트레스에 의한 산화 스트레스 및 항산화 반응의 통합 네트워크는 Fig. 3에 제시하였다.
저온 스트레스로 인한 에너지 분배 재편, 지질대사 변화, 산화 스트레스 및 HSP 반응은 상호 긴밀하게 연결되어 있으며, 이러한 생리적 부담을 완화하기 위해서는 영양적 및 관리적 개입 전략이 필수적이다. 최근 연구들은 저온 스트레스에 대한 대응 전략이 단순한 사육 환경 개선을 넘어, 대사 경로 조절과 세포 보호 기전의 활성화를 표적으로 한 정밀 접근으로 확장되고 있음을 보여준다.
저온 스트레스하에서 지질은 열 생성과 유지 에너지 요구를 충족시키는 핵심 에너지원으로 작용하므로, 지질대사의 효율적 조절은 저온 스트레스 완화 전략의 중심 요소이다. 저온 스트레스는 체온을 유지하기 위해 조류가 더 많은 에너지를 소모하게 함으로써 에너지 요구량을 증가시키며, 그 결과 성장과 생산성에 사용될 자원이 전용된다(Krishnan et al., 2023). 이러한 대사율 증가는 ROS의 생성을 더욱 가속화하여 산화 스트레스를 악화시킨다. 특히 고산지역의 저산소증과 저온 스트레스가 결합되면 상승작용이 발생하여, 조류의 폐고혈압증후군(PHS)에 대한 취약성이 더욱 증가한다(Ahmadipour et al., 2019). 또한 저산소증과 산화 스트레스는 에너지 생산에서 중심적인 역할을 하는 지질대사를 저해한다(Netzer et al., 2015). 지질대사의 핵심 과정 중 하나는 β-산화를 위해 지방산을 미토콘드리아로 수송하는 것으로, 이는 카르니틴 팔미토일전이효소(carnitine palmitoyltransferase, CPT1 및 CPT2)에 의해 매개된다. CPT1은 지방산을 아실카르니틴(acylcarnitine)으로 전환하며, 이렇게 생성된 아실카르니틴은 트랜스로케이스(translocase)에 의해 미토콘드리아 내부로 이동된다. 이후 CPT2에 의해 다시 지방아실-CoA로 전환되어 β-산화와 ATP 생성에 이용된다(Schlaepfer and Joshi, 2020). L-카르니틴은 복강 지방 비율을 감소시키고, 혈중 중성지방, 콜레스테롤 및 LDL 수치를 낮춘다(Azizi-Chekosari et al., 2021). L-카르니틴은 지방산 산화와 에너지 생산을 촉진함으로써 대사 시스템의 부담을 완화한다(Tan et al., 2008). CPT1과 CPT2의 활성을 지원하는 L-카르니틴의 역할은 미토콘드리아 내에서 지방산의 효율적인 수송과 이용을 보장하여, 독성 지질 중간체의 축적을 감소시키고 에너지 항상성을 개선한다. 더 나아가 L-카르니틴의 항산화 특성은 ROS를 중화하여 산화적 손상을 줄이는 데 기여한다(Cetin and Guclu, 2020). Ahmadipour et al.(2025)은 저온 스트레스 조건에서 L-카르니틴 보충이 지방산의 미토콘드리아 내 수송과 산화를 촉진하여 지질대사를 개선하고, 동시에 대사성 질환 발생을 완화한다고 보고하였다. 이러한 결과는 지질대사 조절이 저온 스트레스 적응에 있어 실질적인 효과를 가질 수 있음을 시사한다. 또한, 적절한 사료 에너지 밀도 조절과 지방 공급원 선택은 저온 스트레스하에서 유지 에너지 요구 증가로 인한 성장 저하를 완화하는 데 중요한 역할을 할 수 있다. 이는 에너지 분배 재편이라는 저온 스트레스의 근본적 특성을 고려한 전략적 접근이라 할 수 있다.
저온 스트레스로 증가한 미토콘드리아 ROS 생성은 항산화 방어 능력이 충분히 확보되지 않을 경우 조직 손상으로 이어질 수 있으므로, 항산화 능력 강화는 필수적인 완화 전략이다. 비타민, 미네랄, 그리고 식물 유래 항산화 물질은 ROS 제거 능력을 보완하고 redox balance 유지를 돕는다. 루틴은 식물 유래 폴리페놀로서 초과산화물(superoxide), 과산화물(peroxide) 및 자유 라디칼을 제거할 수 있는 강력한 항산화제이다(Choi et al., 2021; Muvhulawa et al., 2022). 다른 폴리페놀과 비교했을 때, 루틴은 단백질과 지방의 산화적 손상을 예방하는 데 특히 우수한 효과를 보인다(Li et al., 2022). Zhang et al.(2026)은 루틴 보충이 cold-induced oxidative stress를 효과적으로 감소시키고, 항산화 효소 활성 증가와 함께 근육 단백질 구조 안정화를 통해 육질을 개선한다고 보고하였다. 이 연구에서 저온 스트레스를 받은 육계의 흉근에서 Nrf2, HO-1 및 NQO1의 mRNA 발현량이 현저히 감소하였으며, 이는 저온 스트레스가 근육 조직 내 항산화 방어 네트워크를 억제함을 시사한다. 사료에 루틴을 첨가하였을 때 이들 유전자의 발현이 회복되었는데, 이는 루틴이 Nrf2 신호경로를 활성화함으로써 항산화 능력을 증진시킨다는 것을 의미한다. 이러한 항산화 강화 전략은 단순히 산화 손상을 억제하는 데 그치지 않고, 지질대사와 미토콘드리아 기능의 안정화를 통해 저온 스트레스하에서의 전반적인 대사 효율을 향상시킬 수 있다. 따라서 항산화 영양소는 저온 스트레스 완화 전략의 핵심 구성 요소로 고려되어야 한다.
앞선 절에서 논의한 바와 같이, HSPs는 저온 스트레스하에서 ROS buffering과 대사 안정화에 중요한 역할을 수행한다. 최근 연구들은 영양적, 환경적 조절을 통해 HSP 반응을 조절할 수 있는 가능성을 제시하고 있다. 항산화 상태의 개선은 과도한 HSP 유도를 억제하여 세포 에너지 소모를 줄이는 한편, 필요시 HSP 반응이 효과적으로 작동할 수 있는 환경을 조성한다(Balakrishnan et al., 2023). 관리적 측면에서는 간헐적 저온 자극과 같은 전략이 과도한 스트레스 없이 HSP 매개 적응 반응을 유도할 수 있음이 보고되었다. Li et al.(2024)은 조절된 저온 노출이 심장 조직의 에너지 대사를 개선하고 스트레스 저항성을 향상시킨다고 보고하였으며, 이는 HSP 반응을 포함한 세포 보호 기전의 적응적 활성화와 관련될 가능성을 시사한다.
종합적으로, 저온 스트레스 완화 전략은 지질대사 조절을 통한 에너지 공급 안정화, 항산화 능력 강화를 통한 redox balance 유지, 그리고 HSP 반응의 적절한 조절을 통한 세포 보호라는 세 축을 통합적으로 고려해야 한다. 이러한 접근은 단일 영양소 또는 단편적 관리 기법에 의존하는 기존 전략보다, 저온 스트레스의 대사적 본질을 보다 효과적으로 대응할 수 있는 잠재력을 지닌다. 특히 초기 사육 단계에서의 저온 스트레스 관리가 이후 생산성에 장기적 영향을 미칠 수 있음이 보고되고 있어(Moreira et al., 2024), 사육 단계별 맞춤형 영양 및 관리 전략 수립의 필요성이 강조된다. 따라서 저온 스트레스를 단순히 회피해야 할 환경적 위험 요인이 아니라, 관리 가능한 대사적 도전 요인으로 인식하고, 이를 기반으로 한 영양, 사양 관리 전략을 수립하는 것이 향후 가금류 생산성 및 복지 향상에 중요한 의미를 가질 것이다.
결 론
본 총설은 가금류에서 저온 스트레스가 유발하는 생리, 대사 반응을 에너지 분배 재프로그래밍(energy distribution reprogramming)이라는 통합적 관점에서 정리하였다. 저온 스트레스는 유지 에너지 요구량 증가를 출발점으로 열 생성을 강화하고, 이에 따라 지질대사 재편과 미토콘드리아 ROS 생성 증가를 동반한다. 이러한 대사적 변화는 산화 스트레스와 redox imbalance를 유발하지만, 동시에 항산화 방어 시스템과 HSPs의 활성화를 통해 세포 및 조직 수준의 보호 기전을 유도한다. 결과적으로 저온 스트레스 적응은 에너지 대사, 지질대사, 산화 스트레스, HSP 반응이 유기적으로 연결된 통합 네트워크로 이해될 수 있다. 이러한 개념적 틀에도 불구하고, 저온 스트레스에 대한 생리·대사 반응은 실험 조건에 따라 다양한 양상으로 나타날 수 있으며, 이러한 변이성을 통합적으로 이해하는 것이 향후 연구의 중요한 과제가 된다. 가금류에서 저온 스트레스에 대한 이해에는 여전히 여러 연구 공백이 존재한다. 첫째, 대부분의 연구는 단일 조직 또는 단기 노출 조건에 초점을 맞추고 있어, 장기적인 저온 스트레스하에서 조직 간 대사 상호작용을 통합적으로 분석한 연구는 제한적이다. 둘째, 열 스트레스와 달리 저온 스트레스에서는 유전적 요인과 대사 적응 능력 간의 연관성에 대한 연구가 상대적으로 부족하며, Av-UCP와 같은 후보 유전자의 기능적 의미도 충분히 규명되지 않았다. 셋째, 산화스트레스와 HSP 반응이 지질대사 및 생산성 지표와 어떻게 정량적으로 연결되는지에 대한 기전적 증거 역시 제한적이다. 향후 연구에서는 이러한 공백을 해소하기 위해 몇 가지 방향이 제시될 수 있다. 우선, 간, 지방조직, 근육을 아우르는 조직 간 대사 네트워크 분석을 통해 저온 스트레스하 에너지 분배와 지질대사 조절의 전신적 특성을 규명할 필요가 있다. 또한, 유전체․전사체․대사체 분석을 통합한 다중 오믹스 접근은 저온 스트레스 적응과 관련된 핵심 조절 인자와 신호 경로를 규명하는 데 유용할 것이다. 더 나아가, 항산화 영양소, 기능성 사료 원료, 그리고 관리적 저온 자극 전략이 HSP 반응과 대사 안정화에 미치는 장기적 효과를 평가하는 연구가 요구된다. 열 스트레스에 의해 유도되는 지질대사 재프로그래밍에 관한 최근 연구 결과들과 더불어, 저온 스트레스 연구에서 제시되는 새로운 근거들은 가금류가 현저한 대사적 가소성을 보유하고 있으며, 열적 도전 조건에 따라 지질대사가 에너지 보존 또는 에너지 소비 방향으로 재조정될 수 있음을 뒷받침한다.